U potrazi za uzrocima i rešenjima problema gubitaka pčelinjih društava*
Autori:
Stanimirović Zoran - Univerzitet u Beoradu, Fakultet veterinarske medicine, Katedra za biologiju;
Glavinić Uroš Univerzitet u Beoradu, Fakultet veterinarske medicine, Katedra za biologiju;
Ristanić Marko - Univerzitet u Beoradu, Fakultet veterinarske medicine, Katedra za biologiju;
Aleksić Nevenka - Univerzitet u Beoradu, Fakultet veterinarske medicine, Katedra za parazitologiju;
Jovanović Nemanja - Univerzitet u Beoradu, Fakultet veterinarske medicine, Katedra za parazitologiju;
Vejnović Branislav - Univerzitet u Beoradu, Fakultet veterinarske medicine, Katedra za ekomoniku i statistiku;
Stevanović Jevrosima - Univerzitet u Beoradu, Fakultet veterinarske medicine, Katedra za biologiju
* Autorski prevod originalnog rada: Stanimirovic Zoran, Glavinic Uros, Ristanic Marko, Aleksic Nevenka, Jovanovic Nemanja, Vejnovic Branislav and Stevanovic Jevrosima. 2019. Looking for the causes of and solutions to the issue of honey bee colony losses. Acta Veterinaria, 69 (1), pp. 1–31.
Kratak sadržaj: Pčelinji gubici, uključujući one koje su posledica fenomena nestajanja pčelinjih društava (engl. Colony Collapse Disorder — CCD) su i dalje problem savremenog pčelarstva i zbog toga privlače pažnju, kako pčelara, tako i naučne zajednice. CCD karakteriše odsustvo mrtvih odraslih radilica, odnosno pojava da u košnici ostaje samo matica i nekoliko radilica. Legla ima nesrazmerno više u odnosu na broj odraslih pčela, a hrane ima više nego dovoljno. Nema pojave grabeži niti štetočina koje obično napadaju oslabljena društva. U ovom radu su diskutovani uzroci pčelinjih gubitaka, kao i njihova prevencija.
Analizirani su sledeći faktori koji dovode to pojave gubitaka pčelinjih društava: manjak hrane visokog kvaliteta (polen i med), infestacija parazitima, primarno sa Varroa destructor, mešovite virusne infekcije, bakterijske infekcije (Američka i Evropska trulež), gljivične (nozemoza i askosferoza) i tripozomalne infekcije (lotmarioza), nedostatak dobre pčelarske prakse i kvalitetnog menadžmenta na pčelinjaku.
Predlažu se određene preventivne mere: (1) obezbeđivanje visokokvalitetne paše i čiste vode, (2) izbegavanje "šećerizacije" — prekomerne primene šećernog sirupa, (3) zadovoljavanje nutritivnih zahteva pčelinjeg društva, (4) opreznost prilikom prihranjivanja pčela, kako u pogledu vremena, tako u pogledu sastava hrane koja se daje, uz izbegavanje čistog šećernog sirupa koji u prevelikim količinama može dovesti do pojave oksidativnog i energetskog stresa, (5) u slučaju nedostatka prirodne hrane — meda u plodištu, koristiti šećerni sirup sa prirodnim/veštačkim suplementima da bi se izbegla "proteinska glad", (6) obavezna planski organizovana kontrola pčelinjeg krpelja V. destructor zbog njegove vektorske uloge, (7) poštovanje higijensko-sanitarnih mera, kao i principa dobre pčelarske prakse i upravljanja pčelinjacima.
U zaključku, sve preventivne mere treba sprovoditi u skladu sa zakonskom regulativom koja se odnosi na redovnu prolećnu i jesenju kontrolu zdravlja pčela. Navedene mere obavljaju licencirani doktori veterinarske medicine koji, ukoliko je potrebno, predlažu adekvatne tretmane.
Ključne reči: Apis mellifera, gubici pčelinjih društava, ishrana, patogeni medonosne pčele, pesticidi
Uvod
Gajene medonosne pčele (Apis mellifera) imaju mnogo veći značaj zbog uloge u oprašivanju biljaka, nego zbog proizvodnje meda i drugih pčelinjih proizvoda [1, 2]. Stoga je razumljivo da sve veći gubici pčelinjih društava u SAD i Evropi, zabrinjavaju istraživače i pčelare, posebno zbog toga što još uvek nema saglasnosti o konkretnom uzroku (ili uzrocima) gubitaka. Opsežna istraživanja su dokazala da ne postoji pojedinačni faktor koji se može "optužiti" za gubitke, te se zaključuje da je uključen veći broj stresora: nedostatak adekvatne paše, prisustvo patogena, parazita, agropesticida, ali i greške u pčelarskoj praksi [3–10]. Stoga, je teško predložiti samo jedno opšte i najefikasnije rešenje [11]. Podaci iz prošlosti ukazuju da je i ranije dolazilo do pojave visokih gubitaka, ali njima nije pridavan veći značaj. Iako se veliki gubici pčelinjih društava dešavaju širom sveta, njihov uzrok varira. Sprovedena su brojna međunarodna istraživanja sa ciljem utvrđivanja obimnosti i uzroka pčelinjih gubitaka, od kojih ovde prikazujemo rezultate novijeg datuma od dve nezavisne grupe naučnika. Dvogodišnji monitoring 6 000 košnica u 17 zemalja Evrope dokazao je da procenat zimskih gubitaka varira u opsegu od 2 do 32 procenta, kao i da su gubici često sezonski, u skladu sa sezonskom dinamikom životnog ciklusa patogena [12]. Monitoring gubitaka tokom zime 2016/2017. spoveden putem COLOSS ankete u kojoj je učestvovalo 14 813 pčelara iz 27 Evropskih država, Alžira, Izraela i Meksika, dokazao je da je od ukupno 425 762 društava, 5,1 procenata imalo nerešive probleme sa maticom, a 14,1 procenata nije uspelo da preživi zimu [13]. Druge prirodne nepogode su dodatno doprinele gubitku 1,6 procenata od ukupnog broja pčelinjih društava, što kad se sabere ukupno iznosi 20,9 procenata pčelinjih gubitaka. Sami gubici su varirali između država i bili su drastično veći kod pčelara koji imaju mali broj društava. Analiza je dokazala da selidbeni tip pčelarenja nije značajno uticao na zimske gubitke, ali da je imao izvestan negativni uticaj u pojedinim državama [13].
Zbog postojanja većeg broja faktora koji dovode do gubitaka pčela, komplikovano je sprovesti precizna istraživanja u kojima se procenjuje uticaj svakog faktora pojedinačno [6]. Uprkos naglom padu broja gajenih pčelinjih društava u Evropi (25 procenata u centralnoj Evropi u periodu 1985–2005) i Severnoj Americi (59 procenata u periodu 1947–2005), na svetskom nivou je broj pčelinjih društava porastao za 45 procenata (u periodu 1961–2008), zahvaljujući povećanju broja pčelara u nekim državama, pre svega Kini i Argentini [6]. Zbog toga se smatra, da osim zimskih gubitaka, treba posmatrati broj i gubitke pčelinjih zajednica tokom cele godine. Zimski gubici ne odražavaju uvek realno stanje ukupnog smanjenja brojnosti društava.
Pčelinje gubitke ne treba poistovećivati sa specifičnim fenomenom koji je poznat kao sindrom nestajanja pčelinjih društava (engl. Colony Collapse Disorder — CCD) [7], koji je prepoznat i opisan od strane van Engelsdorp-a i sar. 2009. [14], kao stanje koje se manifestuje naglim uginućem društva, bez prisustva mrtvih radilica unutar i ispred košnice. Najčešće u košnici ostaje matica sa malim brojem preživelih radilica. Vidljive su veće površine legla, koje su disproporcionalne u odnosu na broj prisutnih radilica. Zanimljivo je i da u košnicama ima dovoljno hrane (meda i perge), a karakteristično je i da do grabeži i invazije od strane štetočina (voskovih moljaca i male košničke bube) dolazi znatno kasnije, nego što bi se očekivalo [11, 14–16].
Prema Evropskoj agenciji za bezbednost hrane — EFSA [17], najčešći uzroci pčelinjih gubitaka su:
- nedostatak visokokvalitetne hrane (polena i meda),
- parazitske infestacije (pre svega uzrokovane sa Varroa destructor) i mešovite virusne infekcije vezane za vektorsku aktivnost V. destructor,
- bakterijske infekcije (Američka i Evropska trulež), gljivične (nozemoza i askosferoza) i druge infekcije i
- menadžment u pčelarskoj proizvodnji.
Nedostatak visokokvalitetne hrane
Globalne klimatske promene, zagađenje životne sredine i "hemizacija" prisutna u svim sferama, a posebno u poljoprivredi, vode do poremećaja u ekosistemu, biljnoj proizvodnji i oskudici visokokvalitetne hrane za pčele. Promene u dinamici cvetanja, kvantitetu i kvalitetu polena i nektara [18], kao i praksa gajenja monokultura (npr. kukuruza) čiji je kvalitet slabiji za pčele, dodatno povećavaju rizik nastanka proteinske gladi kod pčela [19–21]. U proteklih nekoliko decenija, diverzitet medonosnih biljaka se promenio na globalnom nivou. Tako je u Srbiji, beli bosiljak (Stachys annua) potpuno iščezao, a kod nekih drugih biljaka došlo je do pada u proizvodnji polena i nektara, kao kod bele deteline (Trifolium repens), crvene deteline (T. pratense), matičnjaka (Melissa officinalis), timijana (Thymus serpyllum) i nane (Mentha piperita) [22]. Intezivna upotreba pesticida takođe negativno utiče na proizvodju polena koga pčele transformišu u kvalitetnu hranu — pergu, koja je glavni izvor proteina za pčele starosti od 3 do 18 dana i za starije otvoreno leglo [18, 22, 23]. Dalje, smanjenje broja pašnih životinja (najviše ovaca i koza), a samim tim i manja količina prirodnog đubriva, dovode do smanjenja plodnosti zemljišta i značajno slabijeg razmnožavanja medonosnih biljaka. Kvalitet polena se menja tokom godine. Najkvalitetniji polen daju ranocvetajuće biljke (biljke prolećne paše): leska (Corylus avellana), bela vrba (Salix alba), crna vrba (S. nigra), kukurek (Helleborus odorus), visibaba (Galanthus nivalis), razne vrste ljubičica (Viola spp.), razne vrste kopriva (Lamium spp.), voćke, maslačak (Taraxacum officinale) i trave iz familija Poaceae.
Zanimljivo je da je ukupan, broj košnica u porastu, ponajviše usled potreba pčelara da povećaju prihod, što dovodi do prenaseljenosti određenih teritorija sa košnicama i manjka hrane za pčele [10]. U pokušaju da ostvare veći profit, pčelari ne obraćaju pažnju na potrebe pčela: uzimaju im med čak i iz plodišta, a taj med pripada isključivo pčelama jer predstavlja idealno balansiranu, visoko energetsko-proteinsku hranu koja je najvažniji faktor za uspešno prezimljavanje pčela i brz prolećni razvoj pčelinjeg društva. Mešanjem meda iz plodišta i medišta pčelari čine štetu i pčelama i sebi. Pčele ostaju bez najboljeg izvora energije i visoko-proteinske hrane, koju pčelari pokušavaju da nadomeste dajući pčelama šećerni sirup. Tim postupkom, oni pčelama obezbeđuju samo dodatni izvor energije, a negativno deluju na razvoj i preživljavanje društva, remete strukturu zimskog klubeta, izazivaju energetski i oksidativni stres, pad imuniteta i stimulišu razvoj i umnožavanje patogena [14, 18, 20, 24–31]. Sve navedeno dovodi do poremećaja u podeli rada među pčelama i smanjenja broja ili poptunog odsustva sanitarnih pčela (kućne pčele starosti 12–18 dana) i smanjenja higijensko-negovateljskih aktivnosti [32–39]. Temporalni polietizam (pojava da tip posla zavisi od uzrasta, tako da svaka radilica sukcesivno obavlja čitavu seriju različitih poslova) je od vitalnog značaja za pčelinju zajednicu. Radilice specijalizovane za obavljanje različitih poslova blisko komuniciraju, što osigurava opstanak pčelinjeg društva, ali i lakšu distribuciju patogena u košnici (slika 1).
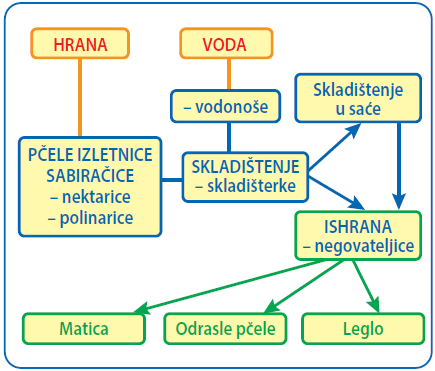
Slika 1. Protok hrane i vode u pčelinjem društvu i komunikacija između različitih kasti pčela
Pored toga, kod neadekvatno hranjenih pčelinjih društava se često zapaža smanjenje broja radilica i trutova, slabija vitalnost i otpornost na patogene (primarno Nosema ceranae) [40]. Osim toga, med uzet iz plodišta može sadržati rezidue različitih preparata (amitraz, kumafos, cimiazol-hidrohlorid, flumetrin, fluvalinat, dicikloheksilamin i fumagilin), koji kontaminiraju med direktno (kada se koriste u jesenjim i zimsko-prolećnim tretmanima), ili indirektno, iz već kontaminiranog voska. Ovaj med nije pogodan za korišćenje u ishrani ljudi i poznato je da rezidue antibiotika i pesticida mogu izazvati razne genotoksične efekte [41–48].
Infestacija sa Varroa destructor i mešovite virusne infekcije.
Pčelinji krpelj Varroa destructor je obligatni parazit za A. mellifera i predstavlja najveću opasnost za pčelarstvo. Usled nedostatka efikasnog načina kontrole ovog parazita, infestirana pčelinja društva uginu za 2 do 3 godine [49]. Zajedno sa pčelinjim virusima koje V. destructor prenosi, ovaj krpelj je jedan od glavnih uzročnika zimskih gubitaka pčelinjih društava [7, 50–53]. U skorije vreme, utvrđeno je da se Varroa hrani masnim tkivom larvi i odraslih pčela [54], a ne isključivo hemolimfom, kako se ranije smatralo [49, 55, 56]. Takođe, reproduktivni potencijal je veći kod krpelja koji jedu masno tkivo u poređenju sa onima koji konzumiraju hemolimfu [54].
V. destructor šteti pčelama direktno, iscrpljujući ih usled remećenja njihovih metaboličkih procesa pre svega metabolizma proteina [57] i indirektno, kao vektor i/ili aktivator pčelinjih virusa [52, 58–60]. Pčelinji krpelj i virus bolesti deformisanih krila (DWV) zajedno ispoljavaju najštetnije dejstvo na pčele jer dovode do skraćenja njihovog životnog veka i spadaju u najčešće uzroke pčelinjih gubitaka na globalnom nivou u poslednjih 50 godina [55, 56, 61, 62]. Međutim, drugi virusi ili njihove kombinacije (virus akutne paralize pčela — ABPV ili komplex ABPV i Kashmir-Israeli virusa — AKI) mogu izazvati pčelinje gubitke zajedno sa Varroa krpeljima kao vektorom koji povećava ukupan titar virusa [51, 52, 63, 64].
Ispitivanjem mehanizama sinergističkog delovanja između V. destructor i DWV, Nazzi i sar. [62] su dokazali da krpelj može poremetiti dinamiku razvoja DWV u telu pčele i indukovati transformaciju virusa u brzo-replikujućeg "ubicu", koji dostiže smrtonosnu virulenciju pri kraju sezone. Kao posledica destabilizacije životnog ciklusa DWV, dolazi do pada aktivnosti imunskog sistema domaćina zbog zanačajnog smanjenja ekspresije faktora NF-kB [62] i poremećaja različitih aspekata imuniteta koje ovaj faktor reguliše. Tu spadaju: sinteza antibakterijskih pepida, agregacija hemocita na mestu povrede i melanizacija, kao i antivirusni mehanizmi odbrane [65]. Smatra se da imunosupresivni efekat, primarno izazvan virusima, povećava negativni uticaj transkripcionog profila više imunskih gena pčela. Takođe, u odsustvu virusa, Varroa ne utiče na ekspresiju dorsal-1A gena, indikatora imunosupresije, za razliku od DWV, koji izaziva najveće smanjenje ekspresije ovog gena [62]. Ova imunosupresivna aktivnost je objašnjena kao deo strategije koju DWV koristi da nadvlada glavne elemente antivirusne zaštite domaćina i time omogući pritajene infekcije. Međutim, bilo koji stresor spoljašnje sredine (pesticidi ili loša ishrana) može poremetiti delikatan, balansirani odnos između virusa i odbrambenih mehanizama pčela, što dovodi do aktivacije odgovora preko NF-kB i intezivne replikacije virusa u pčelama u kojima je infekcija do tada bila u pritajenom obliku (inaparentna infekcija). Najčešća posledica brze replikacije virusa je kolaps pčelinjeg društva [61, 62, 66]. Dalja istraživanja mehanizama interakcije Varroa-virus i pojave pčelinjih gubitaka ukazala su na postojanje simbioze između Varroa parazita i DWV, sa svrhom savladavanja imunskih barijera domaćina [55].
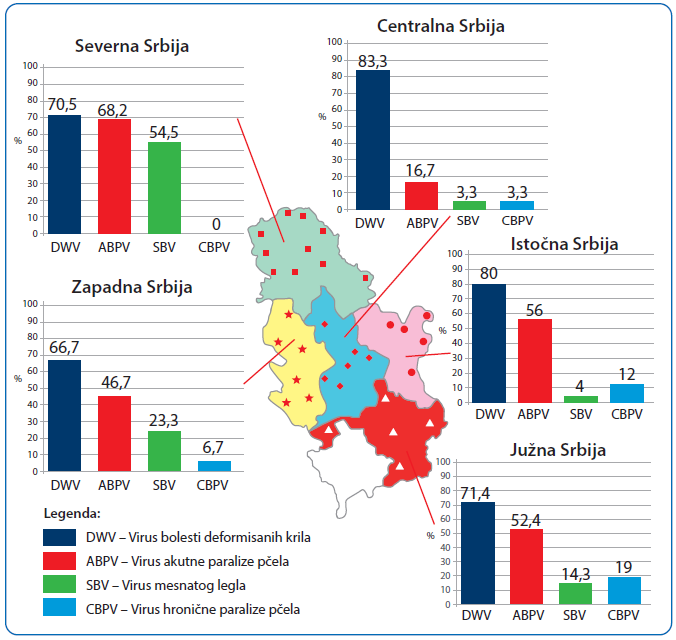
Slika 2. Rasprostranjenost ispitivanih virusa u pčelinjim društvima u pet regiona Srbije [69]
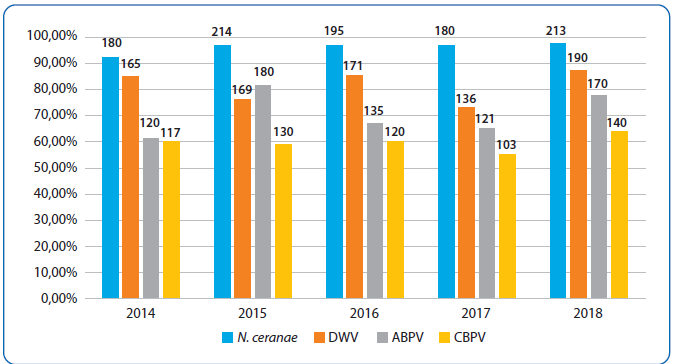
Slika 3. Prisustvo N. ceranae, DWV, ABPV i CBPV kod pčela analiziranih u Laboratoriji za genetiku domaćih životinja, divljači i pčela, Katedre za Biologiju Fakulteta veterinarske medicine Univerziteta u Beogradu (2014–2018)
Dodatna istraživanja imunskog odgovora pčela na simultanu infestaciju Varroa parazitom i infekciju DWV su ukazala da su pčele u stanju da odmah reaguju jakim imunskim i homeostatskim odgovorom, koji ne traje dugo i koga prati remećenje regulacije ovih puteva, čineći pčele ranjivim na izrazitu replikaciju virusa [56]. Fine i sar. [67] su objavili da čak i inertne supstance u agrohemikalijama mogu doprineti nekim negativnim efektima po pčele, uključujući i povišeni mortalitet indukovan virusom. Herbicidi, iako su dizajnirani da spreče rast korova, ograničavaju dostupnost floralnih resursa i negativno utiču, povećavajući nutritivni status pčele, a time i indirektno utiču na ishod virusnih infekcija pčela.
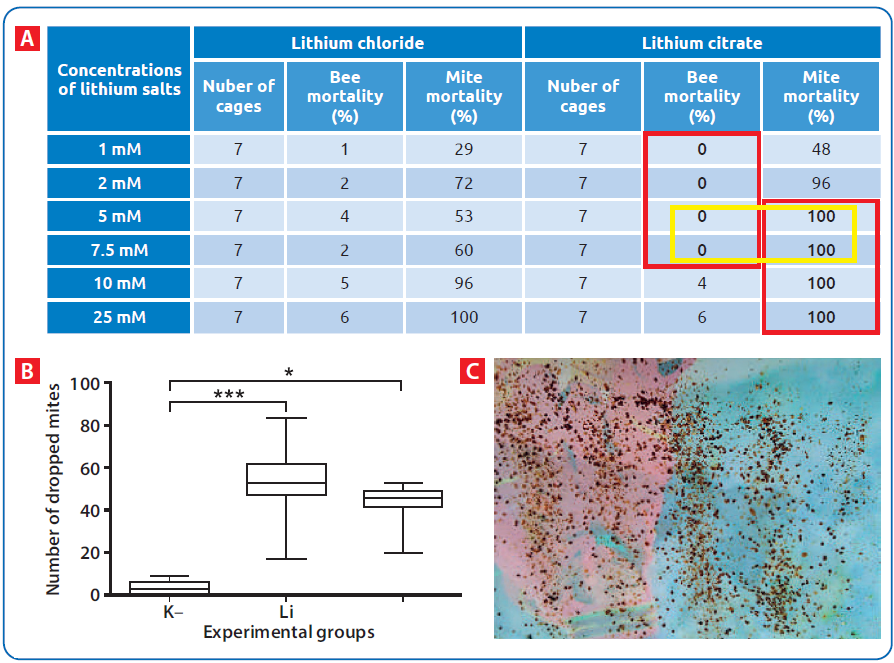
U Srbiji su prva molekularno-genetička ispitivanja prisustva pčelinjih virusa obavljena u našoj laboratoriji [68], a sprovedena su na 11 pčelinjaka i obuhvatila 55 pčelinjih društava iz različitih regiona. Real-time RT-PCR je potvrdio prisustvo DWV na svim pčelinjacima, a ABPV u 10 od 11. Slična, ali opširnija istraživanja smo obavili 2018. godine [69], ispitujući rasprostranjenost (prevalencu) DWV, CBPV, ABPV i SBV unutar društava različite jačine lociranih u 5 regiona Srbije (slika 2).
Analize su potvrdile prisustvo barem jednog virusa u 87,33 procenata pčelinjih društava. Pojedinačne infekcije su dokazane u 28,67 procenata društava (21,33 — DWV, 4,00 — ABPV, 2.67 — SBV i 0,67 procenata — CBPV). Kod većine društava (58,66 procenata) potvrđeno je više od jednog virusa. NaJprevalentniji je bio DWV (74 procenata), praćen sa ABPV (49,30 procenata), SBV (24,00 procenata) i CBPV (6,70 procenata). Filogenetska istraživanja su dokazala da se virusi pčela pronađeni u Srbiji, u 93–99 procenata poklapaju sa virusima deponovanim u svetskoj genskoj banci "GenBank".
Značajan broj pčelara u Srbiji svake godine potražuje usluge od Laboratorije za genetiku domaćih životinja, divljači i pčela (Katedra za Biologiju, Fakultet veterinarske medicine) u vidu analiza uzoraka poreklom od obolelih ili uginulih društava. Kod ovih društava, prevalencija virusa u petogodišnjem periodu (2014–2018) bila je: DWV — 73,12–87,16 procenata, ABPV — 61,54–81,45 i CBPV — 58,82–64,22 procenata (slika 3).
Istorija bolesti, bazirana na tvrdnjama pčelara je najčešće ukazivala na:
(1) neadekvatan tretman protiv varoe, ili potpuno odsustvo tretmana,
(2) oduzimanje velikih količina meda i davanje šećera pčelama i
(3) zazimljavanje pčela na suncokretovom medu (koji je poslednja paša u sezoni i često je kontaminiran pesticidima).
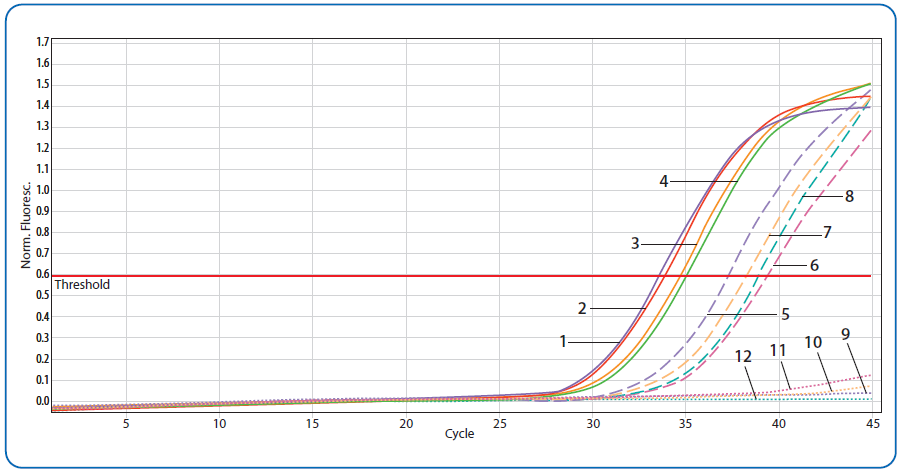
Slika 4. Amplifikacione krive dobijene real-time PCR metodom koje prikazuju detektovane viruse kod odraslih pčela. Linije 1–4 — pozitivne kontrole: 1) Plava linija — ABPV; 2) Crvena linija — CBPV; Narandžasta linija — DWV; Zelena linija — SBV; Isprekidane linije (5–8) — odgovarajući uzorci. Tačkaste linije (9–12) — negativne kontrole. Pojedinačni replikati prikazani su radi bolje preglednosti [70].
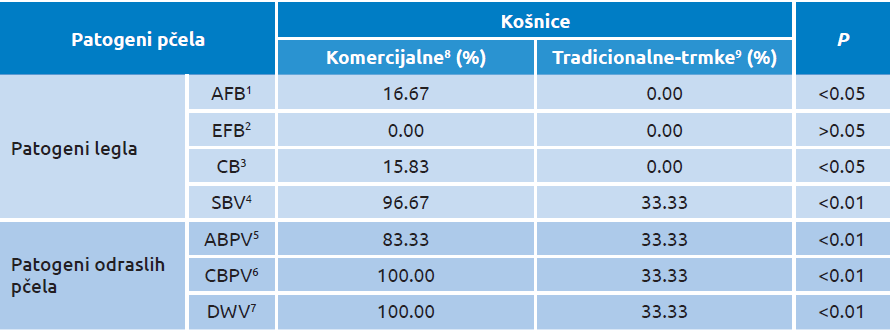
Treba naglasiti da na prisustvo virusne infekcije unutar legla i kod odraslih pčela, utiče tip pčelarenja (konvencionalno-savremeno i tradicionalno). Postoje određeni regioni gde se pčele i dalje uzgajaju na tradicionalni način u primitivnim košnicama načinjenim od vrbovog pruća — takozvanim trmkama. Naša ispitivanja sprovedena na Pešterskoj visoravni, dokazala su da ovakve košnice pružaju znatno bolje uslove za očuvanje zdravlja pčela i njihove otpornosti na patogene [70]. Naizgled zdrava pčelinja društva gajena u komercijalne namene kao i ona koja su gajena u primitivnim košnicama, testirana su na prisustvo virusa mešinastog pčelinjeg legla (SBV), kao i na prisutvo virusa odraslih pčela (slika 4).
U tradicionalnim košnicama SBV je detektovan kod 33,33 procenata, dok je u komercijalnim košnicama utvrđen kod 96,67 procenata uzoraka. Takođe je učestalost virusa odraslih pčela bila značajno veća u komercijalnim košnicama.
Očigledno, kod legla i odraslih pčela odgajanih na tradicionalni način, u primitivnim košnicama, prevalencija svih praćenih virusa bila je do 33,33 procenata bez ispoljavanja kliničkih simptoma, što je u granicama normalne distribucije virusa u pčelinjim društvima u prirodnom uslovima.
Kako bi se sprečili veliki gubici usled mešovitih infekcija varoom i virusima, potrebno je redovno suzbijati varou, vektora različitih patogena [55, 56, 61, 62, 64, 69, 71, 72]. Postoji više načina kontrole V. destructor, a njihova efikasnost je različita. Ove metode su podeljene na biotehničke, hemijske (upotreba sintetskih "hard" akaricida) i ekološke (upotreba prirodnih "soft" akaricida) [49, 73], ali se takođe ove metode mogu koristiti i u kombinaciji [74]. Dugogodišnji rad Stanimirovića i sar. [75] rezultirao je razvojem strategije kontrole varoe koja je primenljiva na Balkanu (slika 5).

Slika 5. Kontrola Varroa krpelja — generalna strategija u Srbiji
Tabela 1. Prevalencija uzročnika bolesti pčela u komercijalnim i tradicionalno uzgajanim pčelinjim društvima na osnovu detekcije njihovih nukleinskih kiselina
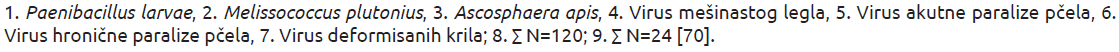
Biotehničke metode oduzimaju mnogo vremena i nisu dovoljno efikasne [74–77]. Upotreba "hard" akaricida, praćena je sledećim problemima: (1) pojava krpelja koji su rezistentni, naročito na piretroide — fluvalinat i flumetrin [78–80] i (2) pojava rezidua u svim proizvodima košnice: najviše koncentracije su dokazane u vosku i propolisu, niže u polenu i pergi, a najniže u medu [49, 79, 81–84]. Najčešće detektovani varoacidi u pčelinjem vosku, polenu i pergi su fluvalinat, kumafos, amitraz, bromopropilat [84–89] i hlorfenvinfos u uzorcima voska i perge iz Španije [90]. S obzirom na to da je vosak sastavni deo košnice, kojem je potrebno najviše vremena za obnavljanje, pesticidi koji ostanu u košnici mogu dovesti do tzv. "sindroma otrovane košnice". Ovaj problem se ne može rešiti zamenom voska jer rezidue lipofilnih akaricida ostaju u pčelinjem vosku čak i nakon recikliranja [89]. Konačno, sintetički akaricidi mogu biti štetni po pčele i uticati na njihove reproduktivne osobine i ponašanje ukoliko se ne koriste ispravno [43, 46, 48, 91, 92].
Radi prevazilaženja svih ustanovljenih problema koji potiču od upotrebe sintetskih akaricida, počela je široka primena akaricida na bazi prirodnih proizvoda, kao što su organske kiseline i biljni ekstrakti [49, 73, 93]. Njihova efikasnost se ispituje već više od dve decenije [73, 93–96]. Mravlja, oksalna i mlečna kiselina su organske kiseline (OA) koje se koriste za suzbijanje varoe. Među biljnim ekstraktima, eterična ulja (EO) i njihove komponente su ubedljivo najviše proučavani kada se govori o varocidnoj aktivnosti [75, 84]. Opšte prednosti prirodnih jedinjenja, OA i EO, su mali rizik od ostavljanja ostataka (rezidua) i nakupljanja u pčelinjim proizvodima, kao i mala verovatnoća razvoja rezistencije nakon ponovljenih tretmana [49]. Ako se pravilno primenjuju, nalaz njihovih rezidua je nizak, pa "imidž" meda i ostalih pčelinjih proizvoda, kao prirodnih, zdravih i čistih, ostaje "neokaljan". Međutim, OA i EO imaju i značajne nedostatke: njihova efikasnost je nedovoljna i zavisi od klimatskih uslova, uslova u košnici kao i načina primene [49, 75, 93]. Neki prirodni preparati imaju neželjene efekte na pčele i/ili njihovo leglo [95, 97]. Srećom, nedavno su stvorene neke formulacije biljnog porekla koje su prilično efikasne, bez neželjenih efekata na razvoj i produktivnost društava [73, 96]. Dakle, nijedna metoda suzbijanja varoa krpelja, koja je korišćena i ispitivana do sada, ne može ispuniti sve ove kriterijume: da bude bezbedna po pčele, da ima visoku efikasnost protiv varoe i da bude jednostavna za primenu. Međutim, uspešna kontrola varoe je ostvariva ako se radi u saglasnosti sa propisima koji se odnose na redovan prolećni i jesenji zdravstveni pregled pčela od strane licenciranih veterinara koji mogu da predlože adekvatne tretmane.
Nedavno je u naučnim krugovima promovisana ideja po kojoj se različita jedinjenja litijuma mogu koristiti u kontroli varoe. Ispitivanje koje su sproveli Ziegelmann i sar. [98] ukazuje da litijumove soli mogu imati izrazit akaricidni efekat. Naše nedavno istraživanje je dokazalo efikasnost suplemenata na bazi litijumovih soli (slika 6A), naročito onih koji sadrže litijum–citrat u različitim koncentracijama. Rezultati su ukazali da koncentracije od 5 i 7,5 mM ispoljavaju snažan akaricidni efekat, bez uticaja na mortalitet (slika 6A). U terenskim eksperimentima, ispitivana koncentracija (7,5 mM) imala je zadovoljavajuću akaricidnu efikasnost (slike 6B i 6C).
Bakterijske infekcije (američka i evropska trulež pčelinjeg legla) i gljivične bolesti (askosferoza i nozemoza)
Američka i evropska trulež pčelinjeg legla su bakterijske bolesti od posebnog značaja u pčelarstvu [99]. Pored ovih bakterijskih bolesti, treba uzeti u obzir invazivnu i destruktivnu mikozu izazvanu sa Ascosphaera apis, kao faktor koji doprinosi slabljenju zdravlja pčela što može voditi njihovom gubitku [100]. Kosmopolitske gljivice Aspergillus spp, zaslužuju veću pažnju zbog visoke virulencije prema larvama pčela i zbog sveprisutnosti njihovih spora u životnoj sredini [101].
Američka trulež legla (AFB) se smatra fatalnom bolešću pčelinjeg legla [50]. U nekim zemljama je česta (npr. utvrđeno je da je 5–10 procenata pčelinjih društava u Nemačkoj zaraženo, bez ispoljenih simptoma) i uzrokuje znatne ekonomske gubitke u pčelarskoj industriji. Klinički znaci i tok AFB variraju u zavisnosti od genotipa Paenibacillus larvae, jačine i bihejvioralnih odbrambenih mehanizama samih pčelinjih društava. Identifikacija genotipova P. larvae (koji se utvrđuju pomoću PCR tehnike) je važna zbog razlika u virulenciji i prognozi bolesti. Genotipovi ERIC I i II ne ubijaju larve u ranoj fazi, nego tek nakon zatvaranja ćelija i zbog toga pčele higijeničarke ne mogu efikasno da očiste bolesno leglo pa dolazi do izbijanja bolesti, a klinički simptomi postaju vidljivi [102]. Genotipovi ERIC III i IV su visoko virulentni za larve i većina larvi ugine pre nego što ćelije budu poklopljene i pčele higijeničarke očiste ostatke, tako da klinički simptomi uglavnom izostaju. Ovi genotipovi P. larvae se razlikuju u klijavosti, otpornosti na različite temperaturne tretmane i periodu očuvanja vijabilnosti nakon skladištenja u zamrzivačima, zbog čega bi uvek trebalo uraditi njihovu genotipizaciju, kao deo standardnih laboratorijskih protokola [103].
Evropska trulež legla (EFB) postoji u pčelinjim društvima širom sveta, uzrokuje ozbiljne gubitke legla i kolaps pčelinjih zajednica [104]. U Švajcarskoj i Velikoj Britaniji, EFB je dovela do velikih problema [105, 106] zbog neuspelih sanitarnih mera, što je ukazalo na neophodnost rešavanja problema širenja M. plutonius [50]. Pored klasičnih mikrobioloških metoda detekcije, za potvrđivanje M. plutonius su razvijene molekularne metode na bazi PCR tehnike [70, 106, 107].
Bolest krečnog legla se često zanemaruje kada se traže uzroci gubitka pčelinjeg društva, uglavnom zbog činjenice da se prisustvo A. apis lako prepoznaje i infekcija lako dijagnostikuje [108]. A. apis uzrokuje visoku smrtnost pčelinjeg legla kao i značajan pad brojnosti pčela u košnici. Jaka društva rešavaju problem regrutovanjem velikog broja pčela higijeničarki koje lako prepoznaju i uklanjaju "kalcifikovane" larve, pa se većina ovih društava oporavi u kratkom vremenskom periodu [109]. Međutim, bolest krečnog legla može doprineti gubitku društava, posebno onih koja su kompromitovana od strane različitih etioloških agenasa. Pojava bolesti takođe zavisi i od tehnike pčelarenja [12, 70].
Slika 6. A) Antivarozni efekat različitih koncentracija litijum citrata i litijum hlorida u kaveznom eksperimentu; B) Antivarozni efekat 7.5 mM litijum citrata. K– Negativna kontrola (netretirana); Li — Grupa tretirana sa litijum citratom; K+ Pozitivna kontrola (tretman sa amitrazom); C) Varroa krpelji otpali pod dejstvom litijum citrata.
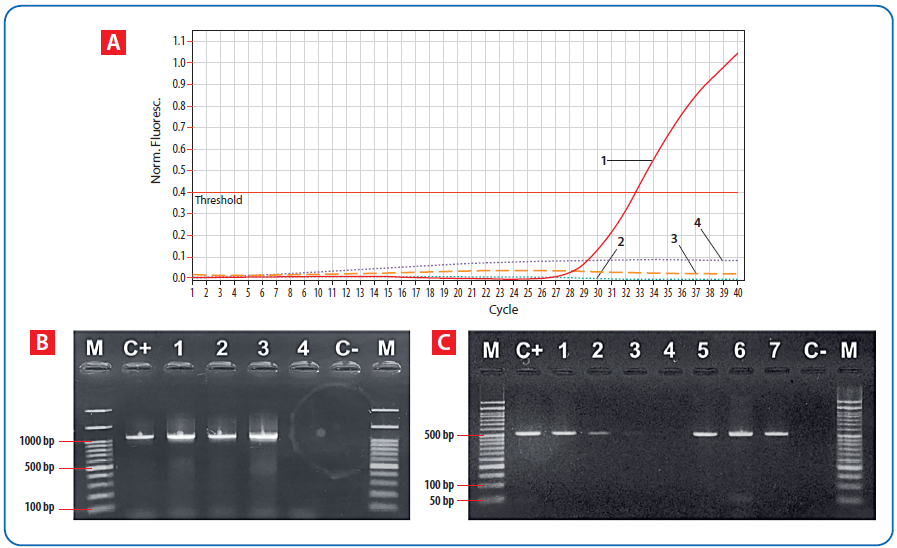
Slika 7. A) Amplifikacione krive dobijene real-time PCR metodom koje prikazuju detekciju M. plutonius u uzorcima legla. Neisprekidana linija (1) – pozitivna kontrola za M. plutonius; Tačkaste linije (2 i 4) – uzorci; Isprekidana linija (3) – negativna kontrola [70]; B) Vizuelizacija produkata amplifikacije, izolata P. larvae. M – 100bp DNK marker; C+ pozitivna kontrola; 1–4 – uzorci; C – negativna kontrola. Veličine pozitivnih traka naznačene su sa leve strane [70]; C) Gel elektroforeza produkata amplifikacije DNK izolata gljivice A. apis. M, 50bp DNK marker; C+, pozitivna kontrola, C – negativna kontrola; (1–7) uzorci. Veličine pozitivnih traka naznačene su sa leve strane [70].
Nedavno smo sproveli istraživanje na Pešterskoj visoravni u Srbiji [70] na naizgled zdravim društvima, čuvanim na tradicionalan način u primitivnim košnicama i onim koja su čuvana u komercijalnim košnicama. Društva su analizirana na prisustvo P. larvae, M. plutonius i A. apis (slika 7). Rezultati su ukazali da tradicionalne košnice pružaju znatno bolje uslove za održavanje zdravlja pčela i njihove otpornosti na patogene: nisu imale bakterijske ili gljivične uzročnike bolesti legla. Suprotno tome, u komercijalnim društvima otkriveni su P. larvae (16,67 procenata), A. apis (15,83 procenata), dok M. plutonius nije pronađen. Dve trećine tradicionalno čuvanih društava bilo je bez ijednog patogena pčela, dok među komercijalno gajenim društvima nije zabeleženo nijedno društvo slobodno od svih analiziranih patogena.
Nozemoza
Mikrosporidije i tripanozomatide su najbrojniji eukariotski crevni paraziti medonosnih pčela [110–114] i povezani su sa povećanim gubicima društava iako je njihova uloga i dalje kontroverzna [115–118]. Postoje tri vrste mikrosporidija koje mogu inficirati A. mellifera: Nosema ceranae, najčešće prisutna i globalno rasprostranjena [116, 119], Nosema apis koja preovladava nad N. ceranae samo u regionima sa hladnom klimom [120–122] i N. neumanni, nedavno opisana, endemska vrsta u Ugandi, koja izaziva slabe infekcije [123]. Smatra se da je N. ceranae ozbiljna pretnja za pčelarsku industriju, ali su samo u nekim regionima, dramatični gubici pčelinjih društava bili jasno povezani samo sa infekcijom N. ceranae [124–126]. N. ceranae kao jedini stresor je ispoljio imunosupresivne efekte dalujući na gene značajne za imunitet u laboratorijskim eksperimentima [127–129]. Međutim, u svim fazama istraživanja postojala je vremenski zavisna nedoslednost u ekspresiji gena (slika 8).
Putem molekularno-genetičkih metoda naša istraživanja su dokazala da N. ceranae predstavlja dominantnu mikrosporidiju medonosnih pčela u Srbiji [110, 111]. Među svim uzorcima koji su sakupljani počev od 2000. godine, samo je za jedan uzorak (poreklom iz 2008. godine) dokazana infekcija sa N. apis, dok su svi ostali bili pozitivni samo na N. ceranae [110, 111]. Prema tome, jasno je da je N. ceranae bila prisutna još od 2000 godine. Zbog odsustva molekularno-genetičkih dokaza da je N. apis ikad bila prevalentna u Srbiji, nema nikakve osnove ni za diskusiju da se desila zamena N. apis sa N. ceranae.
Što se tiče prevalencije N. ceranae u pčelinjim zajednicama u Srbiji, istraživanja Stevanović i sar. (111) su dokazala da je ona kontinuirano visoka jer se kretala u opsegu od 73 do 98 procenata (u periodu 2008–2012); najviša prevalencija je zabeležena u martu i to u opsegu od 94 (2008) do 98 procenata (2010). Niža, ali opet značajna prevalencija zabeležena je u oktobru (67–87 procenata) i junu (73–91 procenata). Ipak, karakteristike nozemoze tip C uzrokovane sa N. ceranae [130] u Španiji (nedostatak sezonalnosti, odsustvo bilo kakvog jasnog simptoma i neminovni kolaps zajednica ukoliko nisu tretirane) nikad nije zabeležen kod N. ceranae-inficiranih pčela u Srbiji [111]. Zapravo je (1) utvrđena sezonalnost u pojavljivanju N. ceranae u periodu 2008–2011; (2) u većini N. ceranae-inficiranih zajednica uočeni su simptomi koji se tradicionalno pripisuju infekciji sa N. apis (mrlje fecesa, mrtve i bolesne pčele koje puze); (3) odsustvo jasne povezanosti između N. ceranae infekcije i gubitaka zajednica ne samo tokom zimske, već i tokom letnje sezone [111]. Kako su simptomi nozemoze uočavani i kod društava koja su kasnije uginula i kod preživelih društava (kod njih čak i češće u periodu 2010–2012) ti simptomi se ne mogu koristiti kao prediktivni markeri, odnosno ne mogu biti merodavni za predviđanje gubitaka pčelinjih društava. Isto važi i za neka druga stanja zabeležena u N. ceranae-inficiranim košnicama: A) rastresito klube tokom hladnih zimskih dana (na temperaturama ispod –5°C) nasuprot kompaktnom kod neinficiranih i B) i povećana uznemirenost pčela tokom nižih temperatura (0°C ili nešto iznad) manifestovana neobično velikim brojem pčela na ulazu u košnicu nakon uznemiravanja [111]. U prilog navedenom ide i nalaz iz naše laboratorije po kome je prevalencija N. ceranae u odraslim pčelama sakupljenim iz košnica sa vidljivim simptomima bolesti tokom petogodišnjeg perioda (2014–2018) bila 92,31–97,71 procenata (slika 3). Prema tome, N. ceranae infekcija pčela u Srbiji, u odsustvu drugih stresora, ne liči na nozemozu tipa C i nije znak koji nagoveštava gubitke pčelinjih zajednica.
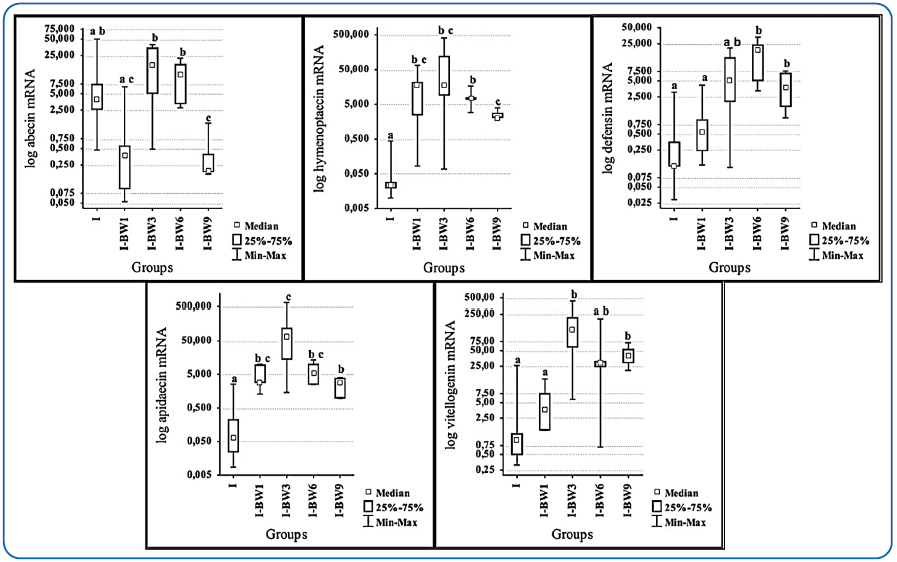
Slika 8. Nivoi ekspresije gena za abecin, himenoptecin, defenzin, apidecin i vitelogenin 12. dana nakon infekcije sporama N. ceranae u grupama tretiranim preparatom "BEEWELL AminoPlus". Pčele u tim grupama su zaražene sporama N. ceranae 3. dana od izleganja i tretirane sa "BEEWELL AminoPlus" prvog (I-BW1), trećeg (I-BW3), šestog (I-BW6) i devetog (I-BW9) dana, dok je kontrolna grupa (I) inficirana sporama N. ceranae, ali nije ničim tretirana. Različita slova označavaju razlike među grupama [129].
Tabela 2. Uticaj stepena infekcije N. ceranae na reproduktivne i produktivne parametre u društvima sa maticama različite starosti [131]
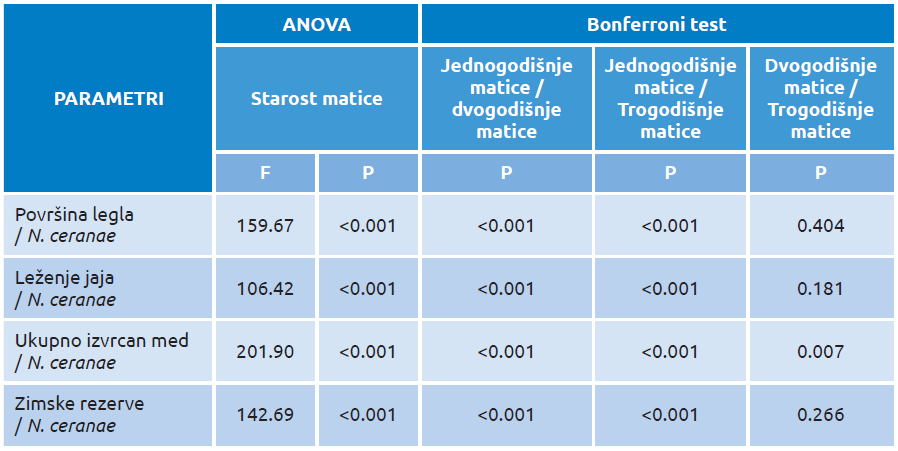
Naša istraživanja su ukazala da N. ceranae može biti uzrok za smanjenje reproduktivnog kapaciteta pčela i proizvodnje meda [131]. Uticaj N. ceranae je ispitan na ujednačenim pčelinjim zajednicama čije su matice različite starosti (1-godišnje, 2-godišnje i 3-godišnje), imajući na umu da bi obnova populacije radilica mogla biti kompromitovana usled isrcrpljenosti matica zbog infekcije sa nozemom. U tim društvima je praćena reproduktivnost, produktivnost i N. ceranae-infekcija (količina spora po zajednici) u trogodišnjem periodu (2009–2012). Značajno veće proizvodne i reproduktivne vredosti zabeležene su u zajednicama sa mlađom maticom (tabela 2). Ovo se može objasniti njihovim većim kapacitetom da kompenzuju gubitke radilica (izazvane infekcijom N. ceranae), a koja se postepeno smanjuje starenjem matice [131].
Ubedljivo najčešće štetne efekte na pčele, ima sinergizam N. ceranae i drugih faktora. Potvrđeno je da istovremeno prisustvo N. ceranae i pčelinjih virusa može da izazove ozbiljne gubitke društava medonosnih pčela [132–134], dok je za kombinaciju N. ceranae i pesticida dokazano da povećavaju smrtnost pčela i menjaju ekspresiju gena povezanih sa imunitetom [135–138]. Međutim, u slučaju kombinacije N. ceranae i tiakloprida, samo su više doze pesticida izazvale značajan mortalitet pčela, zbog toga što je tiakloprid ispoljio negativan uticaj na reprodukciju N. ceranae [139].
Neonikotinoidi su faktori čiji je uticaj na pčele (opstanak, zdravlje, ponašanje, imunitet, reproduktivne i produktivne performanse) najčešće ispitivan. Iako brojna proučavanja nisu omogućila konzistentan zaključak, uglavnom zbog neusaglašenosti između rezultata laboratorijskih i terenskih ispitivanja [140, 141], nedavna istraživanja su pružila dokaze da neonikotinoidi imaju izrazito negativno dejstvo na zdravlje i opstanak medonosnih pčela [142], njihovo ponašanje [143], reproduktivni kapacitet trutova [144] i da u koncentracijama u kojima ih nalazimo na terenu, mogu da oslabe imunološku odbranu [145, 146]. Sveobuhvatne procene rizika izlaganja pčela reziduama pesticida ukazale su da najveći rizik od kontaktne izloženosti reziduama piretroida i neonikotinoida dolazi preko kontaminiranog polena. Šta više, neonikotinoidi predstavljaju mnogo veći rizik u kombinaciji sa EBI (engl. ergosterol biosynthesis-inhibiting) fungicidima zbog njihove sinergističke interakcije [5, 147, 148] ili sa miticidima u košnici — taufluvalinatom, kumafosom i fenpiroksimatom [149]. Jedan od EBI fungicida je prohloraz, široko korišćen u hortikulturi i poljoprivredi, koji je otkriven u medu i polenu uskladištenom u košnicama [150]. Ranije je utvrđeno da on gotovo 1 000 puta povećava toksičnost taufluvalinata, a više od 20 puta toksičnost kumafosa i fenpiroksimata [149]. Prohloraz je, samostalno i u kombinaciji sa kumafosom, promenio ekspresiju gena odgovornih za imunitet kod medonosnih pčela [63, 151]. Nedavno su Glavinić i sar. [138] kod odraslih pčela pratili 15 gena značajnih za imunitet i otkrili da se njihova ekspresija menja kada larve dobijaju hranu koja je kontaminirana prohlorazom (slika 9). Ovi rezultati su dobijeni kombinacijom terenskih i laboratorijskih eksperimenta koji su simulirali uslove u kojima su pčelinja društva (zaražena sa N. ceranae i nezaražena) dovoljno blizu poljima useva tretiranih prohlorazom. U tim uslovima je velika verovatnoća zagađenja useva, te samim tim i legla iz koga će se razviti zimske pčele. Ovaj novi eksperimentalni pristup koji su uveli Glavinić i sar. [138] omogućava uvid u realnu situaciju u kojoj pčelari premeštaju košnice na suncokretovu pašu, koja je često poslednja, pa pčele prezimljavaju na hrani dobijenoj od suncokreta (slika 9).
U suzbijanju infekcije nozemom predlaže se uvođenje dobre pčelarske prakse koja uključuje: zamenu matice novom — mladom, promiskuitetnom i zdravom [152], hranjenje društava visokokvalitetnom hranom (medom i pergom), prihranjivanje šećernim sirupom obogaćenim polenom i/ili suplementima, dezinfekciju košnica i druge opreme, sterilizaciju voska i uklanjanje viška vlage iz košnica. Pored toga, zdrava i bolesna društva nikada ne treba spajati. U nekim zemljama se antibiotik fumagilin i dalje koristi za kontrolu nozemoze. Efikasnost fumagilina je nedavno ponovo procenjena i dokazana u laboratorijskim i terenskim uslovima [153, 154], ali može zavisiti od skladištenja, pripreme za tretman, količine koju pčele konzumiraju i drugih faktora [153]. U Srbiji se, kao i u većem delu sveta, ne preporučuje upotreba fumagilina zbog njegovih štetnih efekata na pčele [154, 155], na kvalitet pčelinjih proizvoda i zdravlje potrošača, koje se narušava prilikom njegove neadekvatne upotrebe i pojave rezidua u medu [44, 45, 47, 156, 157].
Tripanozomne infekcije
Od dva tripanozomatidna parazita medonosne pčele, Crithidia mellificae je poznata otprilike 50 godina i smatrana se široko rasprostranjenom, sve do nedavno kada je opisana Lotmaria passim. Nakon detaljnih genetičkih analiza, ispostavilo se da ova druga preovladava [113]. Neki istraživači [158–162] su ukazivali na mogući uticaj tripanozomatida na zdravlje medonosnih pčela, njihov imunološki odgovor i zimske gubitke društava. Istraživanja patoloških efekata C. mellificae i L. passim na medonosne pčele zahtevaju prvenstveno molekularne alate za identifikaciju ovih tripanozomatida. Stevanović i sar. [112] su dizajnirali i validirali prajmere koji obezbeđuju identifikaciju i diskriminaciju C. mellificae i L. passim putem konvencionalnog PCR ispitivanja, što omogućava rutinsko istraživanje njihove prevalencije i epizootiologije [163]. Osim toga, dizajnirani su i prajmeri za real-time PCR i metoda je optimizovana tako da omogućava istovremenu detekciju i kvantifikaciju L. passim i detaljni monitoring na terenu [114].
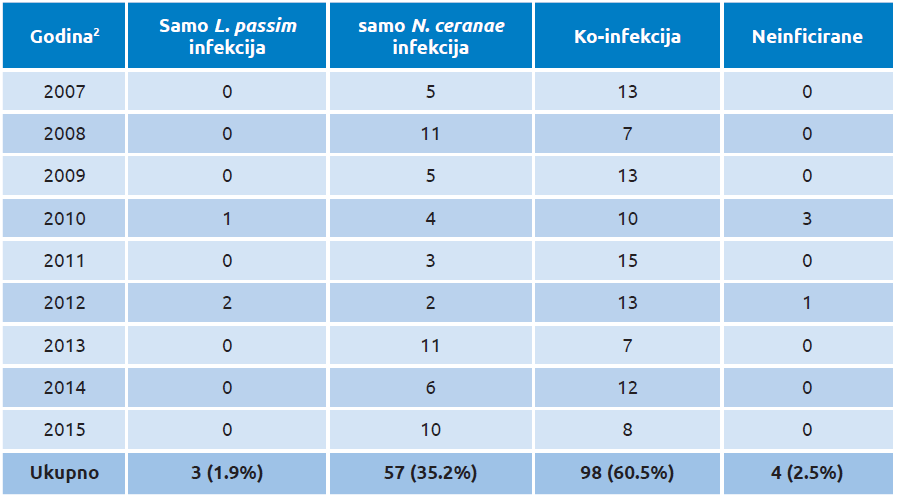
U prvom dugoročnom istraživanju prisustva dve vrste tripanozoma u svetu analizirali smo arhivirane uzorke pčela uzete u Srbiji (2007–2015), ali je otkrivena samo jedna vrsta, L. passim, sa godišnjom prevalencijom od 38,9–83,3 procenata, u proseku 62,3 u devetogodišnjem periodu. Isti uzorci su takođe provereni na prisustvo N. ceranae, koja je pronađena u većini uzoraka sa ukupnom učestalošću od 95,7 procenata i godišnjem opsegu od 83,3 do 100 procenata. Samo je 1,9 procenata pčela bilo zaraženo samo vrstom L. passim, dok su L. passim i N. ceranae istovremeno parazitirale na istom domaćinu u prilično visokom procentu od 60,5 procenata (tabela 3). Štaviše, detektovanje vrste L. passim kod pčela uzorkovanih 2007. godine u Srbiji predstavlja najstariju genetički dokazanu potvrdu ove vrste u svetu i prvu u Srbiji [112].
Visoka pozitivna, statistički zančajna korelacija (r = 0,37; p <0,0001) između nivoa infekcije sa L. passim i N. ceranae ukazala je na njihovu sličnu godišnju dinamiku. Značajne razlike (p <0,05) u nivoima zaraženosti sa obe vrste parazita između meseci ukazuju na sezonski karakter njihove prevalencije. Najveći stepen opterećenosti parazitima N. ceranae i L. passim otkriven je kod pčela izletnica uzorkovanih zimi, a najmanji kod onih uzetih sredinom leta [114].
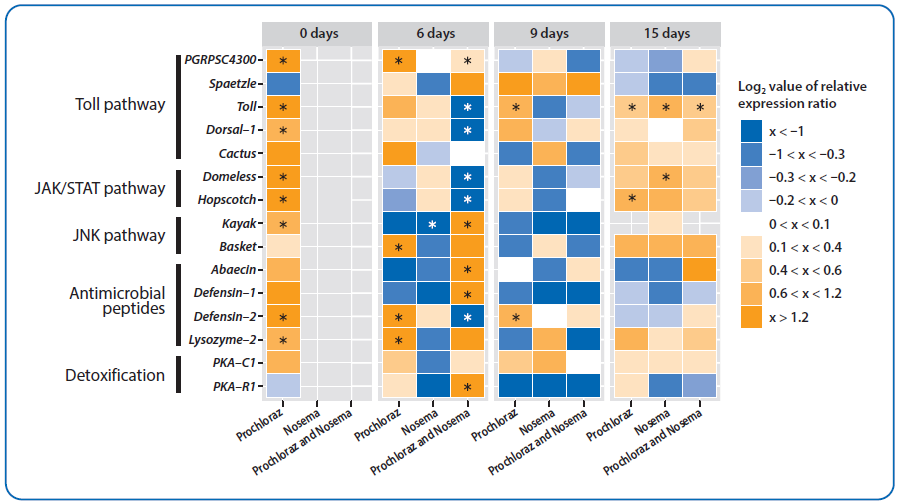
Slika 9. Hitmapa gena značajnih za imunitet kod odraslih pčela različitog uzrasta (0, 6, 9 i 15 dana nakon izleganja). Boje označavaju prosečne nivoe iRNK u poređenju sa prosečnim nivoima iRNK u kontrolnim grupama: plava označava niže, a žuta više nivoe. Vrednost opsega log2 relativnog odnosa ekspresije naznačena je u legendi sa desne strane. Svaki red odgovara transkriptu jednog gena, svaka kolona stepenu ekspresije tokom određenog tretmana. Imena gena i odgovarajući putevi naznačeni su sa leve strane. Tretmani su naznačeni na skali u dnu grafikona (zaraženi nozemom, CN; tretirani prohlorazom, P; tretmani prohlorazom i zaraženi nozemom, PN). Kontrolna grupa (C) je korišćena za normalizaciju. Kvadrati označeni zvezdicom pokazuju statistički značajne efekte tretmana na ekspresiju gena, kada je p-vrednost bila jednaka ili manja od 0,05 [138].
Tabela 3. Godišnji rezime uzorkovanja pčelinjih društava u Srbiji i status infekcije sa Lotmaria passim i/ili Nosema ceranae u period od 2007 do 2015. [112]
Niža aktivnost superoksid dismutaze (SOD), povećane aktivnosti katalaze (CAT) i glutation S-transferaze (GST) i veće koncentracije malonil-dialdehida (MDA) u sezoni, ukazuju na to da infekcija L. passim izaziva oksidativni stres, što može negativno uticati na kondiciju i produktivnost pčela i posledično čini pčelarstvo manje ekonomičnim [164].
Opšti menadžment na pčelinjaku
Pčelarska praksa se danas oslanja na oduzimanju prekomerne količine meda pčelama. Osnovni principi ove ideje potiču iz 60-ih godina prošlog veka u apsolutno različitim uslovima u pogledu klime, raznolikosti flore i agrotehnike. Klima se globalno promenila: u tlu je manje kapilarne vlage usled dugotrajnih perioda suše i smanjenja nivoa podzemnih voda, što je rezultiralo smanjenjem raznolikosti medonosnih biljaka. Pored toga, globalna poljoprivredna hemizacija, kao i hemijske apitehničke mere usmerene na kontrolu pčelinjih patogena dovode do kontaminacije proizvoda u košnici [84, 165, 166]. Treba paziti da se pčelinjaci postave u područja sa dovoljno obilne paše i čiste vode. Blizina paše i broj pčelinjih društava po kvadratnom kilometru površine značajno utiču na zdravlje pčela i na proizvodne i reproduktivne kapacitete pčelinjih društava [23]. Udaljenost između pčelinjaka treba da bude najmanje 1,5 km, kako bi se izbegla konkurencija za sakupljanje hrane, bez obzira na kvalitet dostupne hrane, što znači da treba izbegavati prenaseljenost medonosnih područja košnicama [167].
Tehnička revolucija i razvoj električnih i komunikacionih sistema snažno utiču na orijentaciju pčela u potrazi za hranom i pri povratku u košnice, ali i na razvoj legla i društva u celini. Stoga se mora voditi računa kada se odlučuje gde će se postaviti pčelinjaci i oni treba da budu daleko od nadzemnih dalekovoda i baznih radio stanica [168–170].
Neadekvatno hranjenje pčela šećernim sirupom, kako bi se nadoknadile nedovoljne rezerve hrane u "gladnim godinama", takođe može dovesti do narušavanja kondicije i zdravlja pčelinjih društava i na kraju, do njihovog uginuća. Zbog toga pčelari treba da znaju kako se priprema hrana za pčele i kada je treba dati. Čist šećer je "nužno zlo", a literaturni podaci sugerišu da je mnogo bolje pripremiti i primeniti šećerno-medeni sirup (npr. u odnosu 7 : 3 šećera : meda). Prisustvo meda, koji sadrži razne aktivne komponente, pomaže letnjim pčelama da ga transformišu u nutritivno najkorisniji oblik za pčele. Pčelari treba da znaju da je ovakvo prihranjivanje pčela razumno obaviti na kraju leta (u Srbiji u periodu 1. avgust — 15. septembar), kada ima dovoljno letnjih pčela, koje su jedine sposobne da transformišu sirup u zadovoljavajući oblik za ishranu pčela. Ne bi trebalo izbegavati upotrebu suplemenata u šećernom sirupu, jer se na taj način obezbeđuje dovoljno amino-kiselina, peptida, mikro- i makroelementa kojih nema u čistom šećernom sirupu [18]. Upotreba ovih dodataka može sprečiti energetski, imunski i oksidativni stres kod pčela, a samim tim i gubitke na pčelinjacima [129, 171–174]. Prisustvo mlade, zdrave matice u košnici garantuje razvoj zdravih pčelinjih društava i uspešno pčelarstvo [131, 175]. Odgovarajuća kontrola patogena u košnicama, pre svega pčelinjeg krpelja V. destructor korišćenjem efikasnih, registrovanih varoacida, takođe je preduslov za održavanje pčelinjih društava u dobrom zdravstvenom stanju. Pored toga, istraživanja su potvrdila jasnu vezu između gubitaka pčelinjih društava i obrazovanja, odnosno obučenosti pčelara: profesionalci su bili sposobni da sačuvaju pčelinja društva od bolesti, za razliku od hobista [12, 70]. Profesionalni pčelari su odmah otkrivali simptome, posebno one koji ukazuju na američku kugu ili infestaciju varoom i blagovremeno primenjivali mere suzbijanja, doprinoseći na taj način preživljavanju svojih društava. Ovo su bila istraživanja u kojima je prvi put pažnja bila usmerena na uticaj pčelara i pčelarske prakse na gubitke pčelinjih društava. U istom istraživanju je primećeno da pojava Varroa krpelja, kao "ubice pčela" u Evropu početkom 80-ih godina prošlog veka, nije dovela do povećanh gubitaka pčelinjih društava. To je objašnjeno činjenicom da su pčelari efikasno usvojili mere za borbu protiv ovog krpelja [12].
Zaključak
Postignut je naučni konsenzus da su gubici pčelinjih društava (uključujući i CCD) multifaktorijalni fenomen [3, 4, 6], koji se prema istraživanjima razvija kroz nekoliko faza. Prvo, različiti nespecifični faktori (klimatske promene, agrohemizacija i neadekvatna hrana) smanjuju snagu pčelinjih društava; apitehničke greške (oduzimanje previše meda pčelama i uzastopno dodavanje velikh količina šećera, neadekvatan tretman društava uglavnom protiv V. destructor, visok stres i iscrpljivanje pčela, zazimljavanje društava na medu kontaminiranom pesticidima — suncokretov med i loš trenutak za zazimljavanje društava). Takve pčelinje zajednice postaju lako prijemčive za bakterijske, mikrosporidijalne, tripanozomske i virusne infekcije. Manifestacija nozemoze u kombinaciji sa infekcijom lotmarijom i latentnom američkom truleži pčelinjeg legla, dodatno iscrpljuje pčelinja društva i narušava imunski sistem pčela [176–179]. Konačno, neadekvatne anti-varoa strategije dovode do značajnih zdravstvenih problema kod pčela i širenja virusa kojima je Varroa vektor i/ ili aktivator. To dokazuju i rezultati iz naše laboratorije koji su potvrdili visok nivo virusa u uzorcima obolelih društava praćen velikim opterećenjem spora N. ceranae. Činjenica da je u društvima sa simptomima bolesti otkriveno više spora nego kod uginulih društava, potkrepljuje našu hipotezu da se nakon vrhunca infekcije nozemom (kada je imunitet najslabiji), pčelinji virusi aktiviraju i dolazi do postepenog smanjenja stepena infekcije Nosema parazitom. Virusi, koji su već prisutni u košnicama u Srbiji [68, 69], "čekaju trenutak oslabljenog imuniteta" i postaju krajnji egzekutori pčelinjih društava.
S obzirom na to da je ovo multifaktorijalni problem, preporučena rešenja problema se sastoje od niza aktivnosti:
- Usklađenosti sa dobrom pčelarskom praksom i higijenskim merama, kako na pčelinjaku, tako i u samim košnicama, ali i u objektima u kojima se čuvaju materijali i oprema za pčelarenje i skladište pčelinji proizvodi;
- Postavljanju pčelinjaka na mesta sa obilnom i raznovrsnom medonosnom pašom i čistom vodom;
- Izbegavanju područja u kojima se pesticidi intenzivno koriste, koji mogu, ako su prisutni u nektaru i polenu, ući u košnicu i narušiti zdravlje pčela i ostaviti rezidue u pčelinjim proizvodima;
- Uzgoju zdravih autohtonih pčelinjih društava sa mladim i zdravim maticama i usklađenost sa programima selekcije pčela;
- Obezbeđivanju dovoljnih količina meda, polena i perge za društva tokom zime;
- Preostali med, polen, perga iz košnica koje su pretrpele gubitak društva ne sme se koristiti za ishranu zdravih zajednica. Ako pčele ne obezbede dovoljno hrane, nedostatak treba nadoknaditi medeno-šećernim sirupom ili šećernim sirupom uz dodatak suplemenata;
- Pčelarstvo je danas neodrživo bez upotrebe akaricida za kontrolu Varoa krpelja. Antivaroozne tretmane treba obavljati registrovanim preparatima koji se primenjuju u odgovarajućem periodu godine. Za povećanje efikasnosti mogu se koristiti kombinacije preparata, uzimajući u obzir interakcije između njih. Tako, na primer, u istoj godini kada je korišćen flumetrin ili fluvolinat treba izbegavati upotrebu preparata na bazi kumafosa.
Literatura: Kompletan spisak referenci se može dobiti na lični zahtev.
E mail: mristanic@vet.bg.ac.rs