Značaj mikrobioma u savremenoj živinarskoj proizvodnji [1. deo]
Autori:
Milanko Šekler, Dejan Vidanović, Bojana Tešović, Nikola Vasković, Marko Dmitrić, Kazimir Matović, Zoran Debeljak
Veterinarski specijalistički institut „Kraljevo“, Kraljevo, Srbija
Ove godine, živinsko meso će postati najviše proizvedeno i konzumirano meso na svetu. Očekuje se da će njegova potražnja i dalje rasti, kako bi se zadovoljile potrebe za životinjskim proteinima kod sve veće populacije ljudi. Gastrointestinalni trakt živine je gusto naseljen populacijom mikroorganizama koji blisko i intenzivno stupaju u interakciju sa organizmom domaćina i hranom koja se vari. Mikrobiom creva koristi organizmu domaćina obezbeđujući mu dodatne hranljive sastojke, od inače slabo iskoristivih delova hrane, modulirajući razvoj i funkciju digestivnog trakta i imunskog sistema domaćina. Zauzvrat, organizam domaćina pruža stanište i hranljive materije za rast i razmnožavanje bakterija mikrobioma. Zdravstveni status creva živine ima veliki uticaj na sistemsko zdravlje ptica, dobrobit životinja, efikasnost živinarske proizvodnje, bezbednost hrane i uticaj na životnu sredinu. Skoriji napredak u razvoju tehnologije sekvenciranja (sledeća generacija sekvenciranja), kao metode za identifikaciju i popisivanje svih bakterijskih vrsta prisutnih u nekoj životnoj sredini, probudio je interes za proučavanje uloge mikrobioma creva živine na njihovo zdravlje i produktivnost. Bolje razumevanje funkcije pilećih creva i njihovog mikrobioma pružiće nove mogućnosti za poboljšanje zdravlja živine i živinarske proizvodnje. Značaj ove teme je značajno porastao tokom poslednje dve decenije. Ovaj pregledni rad objedinjuje najnovije informacije o glavnim faktorima koji oblikuju proces sticanja i sazrevanja mikrobioma creva, njihovoj interakciji sa pilećim imunskim sistemom i rezultatima tih interakcija na zdravstveni status creva i produktivnosti jedinki.
Ključne reči: gastrointestinalni trakt, mikrobiološka zajednica creva, mikrobiom, sekvenciranje, živina
"Zelena revolucija" – pravi tvorac savremenog živinarstva
Treba istaći ogroman značaj rezultata koje je iza sebe ostavila uspešno sprovedena "zelena revolucija", čiji je idejni tvorac i pokretač američki agronom Norman Ernest Borloug, koji je upravo za taj svoj doprinos čitavom čovečanstvu dobio Nobelovu nagradu za mir 1970. godine. Pod ovim terminom, "zelena revolucija" je počela u periodu između dva svetska rata (1930. godine), a njeni efekti su postali svima očigledni širom sveta tek u periodu između 1960. i 1980. godine. Ona podrazumeva ogromnu grupu sprovedenih naučnih istraživanja i tehnoloških rešenja koja su bila usmerena na jedinstven naučni cilj, a to je povećanje svetske poljoprivredne proizvodnje i to prvo u najrazvijenijim zemljama sveta. Iz njih je kasnije proizašao nov "praktičan i humanitaran cilj", a to je organizovan transfer "novootkrivenih tehnologija", na ostatak sveta i to pre svega u nerazvijene zemlje i zemlje u razvoju. Krajnji cilj je bio da se svet konačno i zauvek izbori sa problemom gladi, koji je i u tom periodu, kao i tokom čitave istorije čovečanstva, predstavljao stalni Damoklov mač, nad sve brojnijom populacijom ljudi. U tom istom periodu, velika ekspanzija populacije ljudi je bila plod, kako velikih medicinskih otkrića u borbi protiv infektivnih bolesti (otkriće antibiotika, kao i razvoj kvalitetnih vakcina), ali i sociološkopsihološki efekat (prvi duži period stabilnog svetskog mira posle dva velika i razorna svetska rata je naglo povećao natalitet), kao i plod neprestane borbe za potpuno otvaranje svetskog tržišta "rada, kapitala i robe". "Zelena revolucija" je bila najbolji i najočigledniji primer značaja multidisciplinarnog pristupa rešavanju budućih prioritetnih svetskih problema u cilju postizanja trajnih, stabilnih i samoodrživih efekata.
U praktičnom smislu, "zelena revolucija" je predstavljala zajedničke rezultate različitih naučnih istraživanja koji su postignuti u različitim naučnim oblastima: u genetici biljaka (pre svega onih prehrambenih, među njima posebno žitarica: pšenice i kukuruza), u genetici životinja (goveda, svinja, živine), i to kreiranjem hibrida, sorti i rasa koje imaju veliki proizvodni potencijal, zatim razvojem jeftinije i efikasnije infrastrukture za navodnjavanje, ali i snažnim razvojem petrohemije (tehnologije prerade nafte). Ovo je dovelo do nagle ekspanzije proizvodnje veštačkih đubriva, herbicida, pesticida, insekticida, kao i razvoja novih tehnologija za zaštitu zdravlja životinja (vakcina), uz prateću modernizaciju izgradnje tipskih objekata za stočarsku proizvodnju i uz obilje novih zoohigijenskih i zootehničkih mera.
Sve to je zaista imalo za krajni rezultat veliko povećanje svetske poljoprivredne proizvodnje koja je makar "privremeno" obezbedila pristup dovoljnoj količini kvalitetne hrane najvećem delu čovečanstva. Na primer, Indija je sa 10 miliona tona pšenice, koliko je proizvodila 1960. godina, dostigla proizvodnju od 73 miliona već 2006. godine, što je povećanje od skoro 7 puta. U istom periodu, njena populacija je "samo" duplirana, što jasno govori o značaju postignutih rezultata "zelene revolucije".
No, kao što to uvek biva, nijedno dobro ne ide samo i uvek ga prati i neko zlo, koje se obično ne uočava odmah. U ovom smislu, treba istaći da najveći kritičari "zelene revolucije", pre svega, ističu da je drastično povećana proizvodnja hrane "plaćena preskupo" i da je time ugrozila zdravlje čitavog čovečanstva, pre svega zbog upotrebe ogromne količine pesticida i veštačkih đubriva (nezdravih proizvoda petrohemije, to jest prerade nafte), a zatim i zbog narušavanja "dobrobiti životinja". Veoma je važna činjenica da je došlo do preusmeravanja upotrebe ogromnih "viškova" žitarica, umesto za ishranu ljudi, sada za ishranu životinja, što je rezultat novoformiranih "globalističkih i vesternizovanih" navika u ishrani ("kultura brze hrane"), koja je već sama po sebi nezdrava i koja prati "globalizaciju i vesternizaciju" svakog društva ponaosob. Uz to, nagli ekonomski rast velikog broja najmnogoljudnijih zemalja sveta (jugoistočnog regiona Azije) stvorio je ogromne zahteve tamošnjeg, nekada siromašnog seoskog stanovništva (a sada pretvorenog u radničku klasu), za navodno "kvalitetnijom ishranom", za mnogo više "životinjskih proteina", odnosno mesa, mleka i jaja. Takve promene navika u ishrani su u suštini zahtevi za većom stočarskom proizvodnjom, što dalje dovodi do enormnog porasta potreba za novim površinama poljoprivrednog zemljišta. Ovaj resurs je ograničen konačnom površinom planete Zemlje i trenutno se rešava na uštrb krčenja ogromnih površina pod šumama koje su još ostale (na primer Amazona) kako bi se obezbedile potrebne žitarice i biljna proteinska hraniva za ishranu pomenutog ogromnog broja životinja zbog naraslih potreba i zahteva za mesom i tako ponovo u krug. Sve ovo dovodi do drastičnog porasta potreba i za kvalitetnom pijaćom vodom, kao i do posledične proizvodnje ogromne količine đubreta, što je ogroman pritisak na prirodu i za posledicu ima zagađenje kako vodotokova, tako i vazduha.
Oko polovine zagađenja vazduha danas je poreklom od poljoprivredne proizvodnje (pre svega stočarstva), dok celokupna industrija i sve ostale ljudske aktivnosti skupa (uz pridodate uobičajene prirodne izvore zagađenja – na primer vulkani) čine zajedno onu drugu polovinu.
Jasno je da u ovakvoj globalnoj situaciji savremena živinarska proizvodnja danas predstavlja jedan od "najblistavijih" plodova "zelene revolucije". Tako je vreme trajanja tova brojlera skraćeno za preko 2 puta (sa 70 do 80 dana na svega 30 do 40 dana, dok je utrošak hrane po jedinici proizvoda (konverzija) u poslednje 4 decenije takođe impresivno smanjen (sa preko 3 kg na daleko ispod 2 kg). Danas se već postavljaju novi ciljevi u živinarstvu koji predviđaju pad utroška hrane koji će biti sve bliži "čarobnoj jedinici": 1 kilogram koncentrata za 1 kilogram mesa. U poslednjih nekoliko decenija, potražnja za proizvodima živinarstva raste eksponencijalno. Procenjuje se da će u svetu ukupna proizvodnja pilećeg mesa u 2020. godini dostići 130 miliona tona, čime će potrošnja živinskog mesa izbiti na prvo mesto u konkurenciji svih ostalih vrsta mesa (1. OECD/FAO, 2017).
Kao daleki eho gorepomenute "zelene revolucije" država Srbija pamti "zeleni plan", odnosno program za unapređenje poljoprivredne proizvodnje u skladu sa sličnim programima unapređenja koji su se sprovodili svuda u svetu. Dandanas veliki broj "objekata za živinarsku proizvodnju" u Srbiji potiče upravo iz tog perioda kada su rezultati "zelene revolucije" zahvatili ceo svet.
Uvod u mikrobiom i njegovo ispitivanje
Termin mikrobiom je prvi definisao u upotrebio gospodin Džošua Lederberg, dobitnik Nobelove nagrade za medicinu (inače po obrazovanju molekularni biolog koji se bavio mikrobiologijom) i to zbog potrebe, da jednom rečju opiše zbirni genom svih komensalskih, simbiotskih i patogenih mikroorganizama nađenih u ljudskom telu, u gastrointestinalnom traktu, koži, urinarnom i respiratornom sistemu organa (Hooper LV i Gordon JI, 2001). Mikrobiom i mikrobiot se veoma često i u stručnoj literaturi koriste kao da su sinonimi iako nemaju u potpunosti isto značenje. Naime, prema definiciji Koguta (Kogut MH, 2019) mikrobiot predstavlja samo mikroorganizme koji žive u nekoj životnoj sredini (na primer u digestivnom traktu neke životinje) i u sebe uključuje i bakterije, viruse, arheje (protobakterije), gljivice i protozoe, iako se često termin odnosi samo na bakterije. Sa druge strane, termin mikrobiom ima nešto šire značenje i on obuhvata, pored već definisanog mikrobiota (zajednica živih mikroorganizama na jednoj lokaciji), još i prisutne gene, genome, ali i genske proizvode (metabolite i proteine) poreklom i od zajednice mikroorganizama i od domaćina.
Danas ovaj termin najčešće označava zbirni genom svih mikroorganizama kao što su bakterije, arheje, gljivice, protozoe i virusi koji žive u telu nekog domaćina i koji podležu stalnim promenama za vreme čitavog života domaćina (na primer, kao odgovor na domaćinovu ishranu, životnu sredinu, stres, medicinske intervencije i medicinske uslove). Za potrebe ovog rada pod mikrobiomom živine ćemo podrazumevati isključivo bakterije i to pre svega zato što se trenutno mogu naći rezultati istraživanja samo o njima. Podataka o virusima, kao delu mikrobioma živine, uopšte nema. Razlozi su pre svega u tome što je savremena tehnologija detektovanja virusa (sekvenciranje) mnogo zahtevnija i komplikovanija. Ovde možemo samo napomenuti da skoro svaka vrsta bakterija može imati sopstvene viruse koji ih mogu inficirati i lizirati (bakteriofagi). Clavio i Florez (Clavio V i Florez MJV, 2018) u svom radu ističu da je izučavanje bakteriofaga prekinuo pronalazak penicilina, jer su do tada bakteriofagi smatrani kao jedino moguće efikasno sredstvo u borbi protiv bakterija kao izazivača infekcija. Posle otkrića penicilina istraživanja bakteriofaga su nastavljena samo u nekim istočnoevropskim zemljama. Danas su ta istraživanja ponovo u zamahu i već ima prvih registrovanih antibakterijskih preparata (koktel bakteriofaga protiv L. monocytogenes). Prednosti potencijalne terapije bakterijskih infekcija bakteriofagima su ogromne i to pre svega zbog toga što nemaju uticaja na domaćina i njegov ostatak mikrobioma (korisne bakterije), što deluju samo na svoj patogen (bakteriju), a nema ni pojave rezistencije.
Mikrobiom (mikroorganizmi i njihovi genomi koji se nalaze u čoveku ili životinji), koji je do nedavno posmatran samo kao potencijalni izvor patogena za svog domaćina (čoveka i životinje), danas je prepoznat kao kompleksna zajednica mikroorganizama sa veoma važnim uticajem na celokupni zdravstveni status domaćina. Sugestija je da ljude, kao i sve druge višećelijske organizme (i živinu), treba posmatrati samo kao jednu vrstu "supraorganizma" (nadorganizam) koja je međusobno interaktivna sa sopstvenim mikrobiomima (Turnbaugh PJ i sar., 2007). Pored toga, mikrobiom daje značajan doprinos energetskoj homeostazi, metabolizmu, zdravlju, imunološkim aktivnostima i neurorazvoju organizma domaćina (Cho I i sar., 2012).
Građa (sastav) i funkcionalnost mikrobioma creva je krucijalna za zdravlje živine, dok proces nastanka i sazrevanja mikrobioma creva kroz odvijanje razvojnog ciklusa živine ima snažan uticaj na razvoj intestinalnog epitela i modulaciju fizioloških funkcija koje su neophodne za očuvanje intestinalne homeostaze (imunitet, varenje hrane, očuvanost crevne barijere), a što je sve skupa neophodno za optimalnu i efikasnu ekstrakciju, a potom i naknadnu upotrebu izdvojene energije od strane samog domaćina (Kers JG i sar., 2018). Integritet (očuvana celovitost) gastrointestinalnog trakta i mikrobiološka zajednica creva igraju vitalnu ulogu u apsorpciji nutrijenata (hranljivih sastojaka hrane), razvoju imuniteta i rezistencije na zarazne bolesti. Narušavanje mikrobioma (mikrobiološke zajednice) creva može imati štetne efekte na efikasnost iskoristivosti hrane, produktivnost i celokupno zdravlje živine.
Istorijski gledano, skoro čitav jedan vek, izolacija mikroorganizama, bazirana na upotrebi selektivnih podloga je bila jedina metoda izbora koja je korišćena za identifikaciju i karakterizaciju mikrobiološke raznovrsnosti crevne mikroflore životinja, pa i ptica. Klasične metode izolacije mikroorganizama su široko primenjivane i prilikom ispitivanja mikrobioma creva živine. Ove metode su visoko selektivne za kultivativne bakterije pod određenim uslovima (Hugenholtz P i sar., 1998), dok većina bakterija izolacijom na hranljivim podlogama ostaje neotkrivena jer su nekultivativne (Apajalahti J i sar., 2004). Još pre 30 godina je ustanovljeno da samo jedan veoma mali deo, svega 0,1–1 procenata prisutnih vrsta mikroorganizama u nekom uzorku, može biti uzgajan u veštačkim uslovima (u laboratoriji), odnosno izolovan umnožavanjem (Staley JT i Konopka A, 1985).
Još pre desetak godina je primećeno da je, od ukupno 52 kola (phyla) bakterija, za koje se do tada znalo, samo polovina imala bar jednog jedinog predstavnika koji se mogao "kultivisati" (izolovati), podupirući time tezu da postoji ogromna "nekultivabilna većina" (Rappe MS i Giovannoni SJ, 2003). Zbog toga su raznovrsnost i bogatstvo prisutnih vrsta bakterija u mikrobiomu creva redovno bili potcenjeni i ostajali skoro nepoznati (Gong J i sar., 2002).
U poslednjoj dekadi, upotreba tehnologije sekvenciranja, i to samo onih delova bakterijskih gena koji kodiraju sintezu jednog dela bakterijskog ribozoma označenog kao 16S ribozomalna RNK (rRNK), dramatično je unapredio i poboljšao naše razumevanje sastava i diverziteta svih mikrobioma, pa i onog kod živine. Ova moderna i brza tehnika sekvenciranja omogućava brzo dobijanje kompletnog popisa prisutnih bakterija (kao i ostalih prisutnih mikroorganizama) na bilo kojoj lokaciji u digestivnom traktu živine, a ujedno predstavlja i moćan alat koji nam omogućava do sagledamo kompleksnu ulogu ukupnog mikrobioma creva živine. Sekvence gena, koji kodiraju sintezu subjedinice ribozoma označene kao 16S rRNK, imaju krucijalnu ulogu u mikrobiološkoj filogenezi i taksonomskoj analizi bakterija i arheja, dok je fragment 18S rRNK gena važan za identifikaciju eukariota, a nuklearni ribozomski unutrašnji transkripcioni "spacer" region je važan kod taksonomije gljivica (Zdunczyk Z, 2019).
Za razliku od klasične mikrobiologije koja se fokusira na sposobnost bakterija da rastu in vitro, metagenomika je izučavanje genetskih informacija poreklom od mikroflore koja ima za cilj da izvrši karakterizaciju celokupnog mikrobioma, odnosno genome svih prisutnih bakterija, arheja (prabakterija, gljivica, protozoa i virusa koji žive u domaćinu). Metagenomika se pak oslanja na NGS sekvenciranje, modernu tehniku masovnog i paralelnog sekvenciranja, koja omogućava milionima reakcija da budu izvedene u isto vreme i paralelno, kako bi se dekodirali kompletni genomi i to cele zajednice mikroorganizama u mikrobiomu. Metagenomika izučava genetski materijal poreklom iz životne sredine ili nekog dela mikrobioma povezanog sa domaćinom kako bi potom opisala mikrobiološki diverzitet i funkciju (Choi KY i sar., 2015). Ove metagenomske analize podržavaju i taksonomske analize (analizirajući gene fragmenata ribozomalne 16S RNK kod bakterija ili celih genoma bakterija) ili funkcionalne analize kako bi se identifikovali geni koji kodiraju proteine koji učestvuju u regulisanju i kontroli postojećih metaboličkih puteva i ciklusa. Tehnički napredak je, prema tome, doveo do razvoja NGS sekvenciranja, masovnog paralelnog sekvenciranja – modernog pristupa za dekodiranje prisutnih celih mikrobioloških zajednica (mikrobioma) (Thomas T i sar., 2012).
Do kraja 2018. godine je objavljeno preko 20 000 naučnih radova koji su imali kao ključnu reč upravo reč mikrobiom. Ovih 20000 naučnih radova je objavljeno u poslednjih 15 godina i to sledećom dinamikom po petogodišnjim ciklusima: u prvih 5 godina – 106 izveštaja, u drugih 5 godina – 2 724 izveštaja, a u poslednjih 5 godina – 17 357 izveštaja. Ovakva dinamika objavljivanja radova jasno svedoči o ogromnom rastu interesovanja za mikrobiomom. Više od 100 radova je bilo fokusirano na mikrobiom gastrointestinalnog trakta živine, dok se ostatak izveštaja odnosio na različite oblasti i pitanja u vezi s mikrobiomom (Zdunczyk Z, 2019).
Dok se kod klasične mikrobiologije mikroflorom smatra populacija živih mikroorganizama, a rezultati izolacije odnose samo na populaciju živih mikroorganizama (izražene u "kolonija formirajuće jedinice" – CFU), termin mikrobiom se odnosi na zbirni sadržaj genoma mikroflore, gde se ekvivalentom ustanovljene vrste mikroorganizama smatra "operativna taksonomska jedinica" (OTU) koja predstavlja predstavnika određene vrste mikroorganizama ako formira klasterizovanu jedinicu sa visokom sličnošću svih sekvenci, najčešće višom od 97 procenata. Broj OTU jedinica je disproporcionalno veći, jer one obuhvataju mnogo kompleksniju mikrofloru (bakterije, gljivice, viruse, itd.). Pored toga, bakterijski genomi različitih izolata, koji pripadaju istoj vrsti mikroorganizma, obično ispoljavaju znatnu genetsku heterogenost (Nielsen HB i sar., 2014).
Istraživači metagenomike traže odgovore na sledeća pitanja: "Šta je to tamo?", na koje je odgovor filogenetsko profilisanje (taksonomska analiza), i "Šta oni to rade?", koje se odnosi na funkcionalne genetske analize metabolita bakterija (funkcionalne analize) (Chistoserdova L, 2009). Mikrobiologija je doživela revoluciju uvođenjem visokopropusnih (brzih) metagenomskih tehnologija, koje uključuju sekvenciranje i funkcionalnu analizu genetskog materijala, isto kao i velikim napretkom u bioinformatici koja podrazumeva razvoj računarskih metoda za brzu analizu strukture, funkcija i evolucije gena, genoma i proteina (BordaMolina D i sar., 2016).
Gastrointestinalni trakt živine i njegov mikrobiom
Gastrointestinalni trakt živine uključuje u svoj sastav sledeće segmente: voljku, žlezdani želudac, bubac (mišićni želudac), duodenum (dvanaestopalačno crevo), ileum (tanko crevo), cekume (slepa creva), debelo crevo (kolon) i kloaku. Svaki od navedenih segmenata gastrointestinalnog trakta živine ima različite metaboličke funkcije koje samim tim oblikuju, u tom delu prisutnu, zajednicu mikroorganizama, odnosno mikrobiom.
U voljci živine se nalazi od 108 do 109 ćelijskih bakterija po gramu njenog sadržaja i obično dominiraju bakterije iz roda Lactobacillus (Gong J i sar., 2007).
U želucu je koncentracija bakterija slična pomenutoj koncentraciji bakterija u voljci, ali je nivo fermentacija izazvanih bakterijama mnogo niži u želucu nego u voljci jer je pH želuca manji. Većina bakterija u želucu su laktobacili, enterokoke, laktozanegativne enterobakterije, kao i koliformne bakterije.
Kada govorimo o segmentima tankog creva, koncentracija (ili gustina) bakterija je najniža u duodenumu, što je posledica kratkog vremena zadržavanja sadržaja creva u njemu, ali i zbog ulivanja žuči u tom delu creva koje dodatno razređuje sadržaj creva, to jest hrane koja se vari (Shapiro SK i Sarles WB, 1949). Bakterije koje najčešće čine mikrobiom duodenuma živine su klostridije, streptokoke, enterobakterije i laktobacili (Waite DW i Taylor M, 2015).
Sadržaj mikrobioma tankih creva je jedan od najčešće ispitivanih segmenata creva živine. Lu i sar. (Lu J i sar., 2003) su ispitivali mikrobiom ileuma brojlera i detektovali da Lactobacillus čini glavni mikroorganizam u mikrobiomu (oko 70 procenata), praćen bakterijama iz familije Clostridiaceae (11 procenata), Streptococcus (6,5 procenata) i Enterococcus (6,5 procenata).
Tabela 1. Najčešće prisutne bakterije sistematizovane prema važećoj taksonomiji bakterija (phylum – kolo, order – red, family – familija, genus – rod) u gastrointestinalnom taktu živine posmatrano u odnosu na starost i ishranu
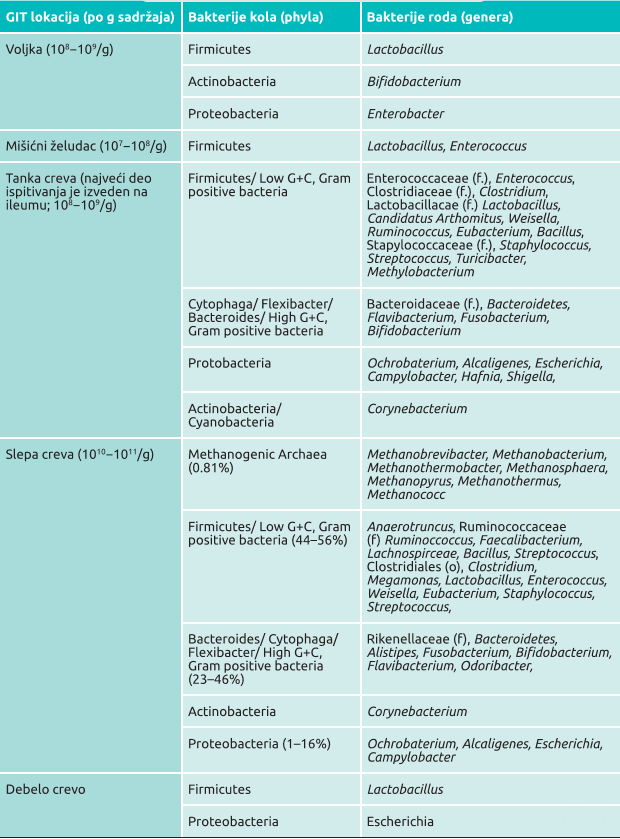
Tabela je modifikovana u odnosu na originalnu verziju (Shang Y i sar., 2018, Chicken Gut Microbiota: Importance and Detection Tech- nology. Frontiers in veterinary science, 5, 254)
U poređenju sa mikrobiomom ileuma, slepa creva živine su imala mnogo raznovrsniji sastav mikrobioma, koji je redovno obuhvatao i anaerobne bakterije (Salanitro JP i sar., 1974; Videnska P i sar., 2013). Oakley i sar. (Oakley BB i sar., 2014) su u svojim istraživanjima dokumentovali značajne promene u sastavu mikrobioma slepih creva posmatrano od prvog dana života pileta pa do starosti od 6 nedelja, ali su isto tako otkrili i značajne razlike u uzorcima poreklom od fecesa i slepih creva, i to posmatrano individualno po pojedinačnim pticama. Do povećanja koncentracije bakterija, kao i njihove sve veće raznovrsnosti u slepim crevima, dolazi tipično u prvih 6 nedelja starosti, dok se taksonomski sastav mikrobioma brzo menja od bakterija Proteobacteria, Bacteroides i Firmicutes do bakterija iz familije Firmicutes, do 3 nedelje starosti (Oakley BB i sar., 2014, Kogut MH i Oakley BB, 2016).
Slepa creva su kao organ digestivnog trakta interesantna sa gledišta mikrobioma jer tamo postoji najveća koncentracija mikroorganizama po gramu (1011), a i hrana se u njima zadržava najduže (od 12 do 20 časova). U slepim crevima se odvija recikliranje uree, regulisanje sadržaja vode, kao i fermentacija ugljenih hidrata. Mada pilići (kojima su eksperimentalno odstranjena slepa creva) mogu da žive i bez njih, ustanovljeno je da se oko 10 procenata energije (od ukupne energije u životinji) obezbeđuje digestivnim procesima u samim cekumima i gde je, baš zbog toga, najveća koncentracija kratkolančanih masnih kiselina (nego u bilo kom drugom delu digestivnog trakta). Firicutes, Bacteroides i Proteobacteria su najvažnija kola bakterija, uz uvek prisutni Actinobacter. Uz to, prisutni su i članovi bakterija Clostridiales za koje se zna da imaju najveći doprinos u produkciji kratkolančanih masnih kiselina. Metagenomskom analizom je ustanovljeno da se u cekumima uvek nalazi velika koncentracija gena povezanih sa metabolizmom ugljenih hidrata koji su poznati iz GenBank baze (metagenomske biblioteke). Takođe je ustanovljeno i prisustvo preko 200 različitih enzima koji učestvuju u razgradnji neskrobnih ugljenih hidrata, od kojih neki upućuju i na potpuno nove metaboličke puteve za proizvodnju propionske kiseline. Takođe je ustanovljeno da hidrogenazu unose bakterije predstavnici rodova Megamonas, Helicobacter i Campylobacter i na taj način omogućavaju proizvodnju kratkolančanih masnih kiselina.
Međutim, Kumar i sar. (Kumar S i sar., 2018) su otkrili da bakterije iz kola Firmicutes predstavljaju najčešće prisutno kolo bakterija i u cekumu i u ileumu u svim uzrastima brojlerske živine (od 1. do 42. dana starosti) izuzev što se u slepim crevima, u 42. danu starosti, beleži i prisustvo bakterija Bacteroidetes. Ove ustanovljene razlike u bakterijskom sastavu mikrobioma mogu poticati i zbog nekih tehničkih razloga kao što su: upotreba različitih protokola za ekstrakciju nukleinskih kiselina, prajmera, protokola sekvenciranja, uslova spoljne sredine gde je smeštena živina, sastava hrane, hibrida živine, kao i geografskih uslova na lokaciji gde se izvode ispitivanja. Različiti rezultati mogu biti i posledica same prirode uzorka koji se ispituje (veća zapreminska količina uzorka voljke nego uzorka slepih creva usled veće količine sadržaja voljke) (Lagkouvardos I i sar., 2017). Međutim, koliko je ovo polje istraživanja složeno, govori i činjenica da su otkrivene velike varijacije u mikrobiološkom sastavu mikrobioma pojedinačnih brojlerskih ptica koje su sve hranjene na isti način i istom hranom, da bi se naknadno otkrilo da je do tih varijacija došlo usled različitog vremenskog perioda koji je protekao od momenta hranjenja do momenta uzorkovanja (Choi JH i sar., 2014).
Način obrade hrane, komponente hrane i aditivi u hrani imaju veliki efekat na mikrobiom creva. Knarreborg i sar. (Knarreborg A i sar., 2002) su ustanovili da koncentrati u praškastom obliku imaju niži procenat bakterija Lactobacillus spp. i C. perfringens u tankim crevima brojlera (ileum), u odnosu na peletirane koncentrate istog sastava. Kukuruz favorizuje manji udeo u mikrobiomu Gram + klostridija, enterokoka i laktobacilusa, dok pšenica favorizuje veći udeo G + bifidobakterija (Apajalahti J i sar., 2004). Gastrointestinalni mikrobiom živine se može podeliti i na mikrobiom koji se nalazi u sadržaju creva i mikrobiom koji se nalazi na mukoznoj površini creva domaćina (Jeurissen SH i sar., 2002).
Sastav mikrobioma lumena creva (ili sadržaja creva) zavisi i od sastava hrane, prisustva antimikrobnih supstanci i brzine kojom hrana pasira crevni trakt domaćina, odnosno živine.
Sastav mikrobioma, vezanog za površinu mukoze creva zavisi od nekoliko faktora i mehanizama domaćina kao što je na primer: ekspresija specifičnih vezivnih (adhezionih) mesta koja se nalaze na površini membrane enterocita, od obima sekrecije imunoglobulina IgA klase, kao i od produkcije mucina. Pored toga, luminalni (feces) i mukozni mikrobiom (površina creva), vrše uticaj jedan na drugi te je zbog toga važno razumeti da ishrana može oštetiti oba pomenuta mikrobioma i dovesti do poremećaja "zdravlja" creva.
Metagenomska analiza sastava intestinalnog mikrobioma creva živine
Pedesetak vrsta bakterija je identifikovano kod jednodnevnih pilića, a taj broj bakterijskih vrsta u sadržaju cekuma se povećava na preko 200 u 42. danu starosti. Udeo mikrobiološke kolonizacije digestivnog trakta je definisan higijenskim standardima, sastavom hrane i unošenjem hrane. Kod tronedeljnih pilića, udeo mlečnokiselinskih bakterija u mikrobiomu ileuma prelazi 70 procenata, dok je udeo drugih grupa bakterija, koje uključuju Clostridium spp., Streptococcus spp., i Enterobacteriaceae spp., bio znatno manji. U završnom stadijuma odgoja primećene su značajne varijacije u sastavu mikrobioma u tankim crevima i cekumima. U kvantitativnom smislu, Lactobacillus bakterije su bile pretežno u ileumu, dok udeo Clostridium bakterija nije prelazio 10 procenata. Sasvim je obrnuta proporcija zastupljenosti ovih bakterija u mikrobiomu cekuma. Odnos Lactobacillusa prema Clostridiumu je bio 7,75 procenata prema 39,3 procenta.
Skorija istraživanja ukazuju da mikrobiom digestivnog trakta živine sadrži više od 500 filotipova bakterija, što čini oko milion različitih bakterijskih gena. To je za 40 do 50 puta više od celokupnog genoma pilića (Oakley BB i sar., 2014). Prema procenama nekih drugih autora, gastrointestinalni mikrobiom živine se sastoji od 640 vrsta bakterija (Apajalahti J i sar., 2004), pa čak sve do 900 identifikovanih vrsta bakterija (Wei S i sar., 2013), od kojih je samo oko 10 procenata moguće izolovati klasičnim bakteriološkim metodama u laboratoriji.
Sekvence identifikovane u digestivnom traktu živine i ljudi predstavljaju 13 bakterijskih kola (phyla) (Wei S i sar., 2013) sa ubedljivim preovladavanjem bakterija iz filuma Firmicutes i Bacterioidetes, od kojih čak 10 predstavlja samo mali udeo mikrobioma i to ispod 5 procenata (Oakley BB i sar., 2014).
Firmicutes bakterije dominiraju svojim prisustvom u gornjim partijama digestivnog trakta (čine oko 90 procenata zajednice mikroba) i predstavljaju ih uglavnom bakterije Lactobacillus, Clostridium i Streptococcus (Wei S i sar., 2013). Jedno drugo ispitivanje navodi da je u crevima udeo bakterija Firmicutes procenjivan na 44–55 procenata, a udeo bakterija Bacteroidetes, predstavljenih rodom Bacteroides je procenjivan između 22–45 procenata, dok su bakterije Actinobacteria (uključujući Bifidobacterium) i Proteobacteria (uključujući Escherichia i Shigella) bile daleko manje prisutne – ispod jednog procenta (Qu A i sar., 2008). U studiji koju su izveli De Cesare i sar. (De Cesare A i sar., 2017) u cekalnom sadržaju su bile pretežno bakterije Firmicutes (93,9 procenta), dok je udeo Proteobacteria bio 1,74 procenta, Actinobacteria 0,92 procenta, Bacteroidetes 0,35 procenata i Tenericutes 1,18 procenata.
Wei S i sar. (Wei S i sar., 2013) su poredili mikrobiom pilića i ćuraka. Mikrobiom ćuraka je bio manje diverzifikovan od mikrobioma pilića, ali su kod obe vrste živine, bakterije Firmicutes, Bacteroidetes i Proteobacteria bile najbrojnije i činile su preko 90 procenata svih detektovanih sekvenci bakterija. Pretežno prisutni rodovi kod pilića i ćuraka su bili Clostridium, Rumnicoccus, Lactobacillus i Bacteroidetes, ali se njihova distribucija po lokacijama u digestivnom traktu umnogome razlikovala. U studiji Singa i sar. (Singh KM i sar., 2014) kod pilića sa visokom konverzijom hrane (engl. feed conversion ratio – FCR) je bilo karakteristično češće prisustvo bakterija iz rodova Acinetobacter, Bacteroides, Streptococcus, Clostridium i Lactobacillus, dok su kod ptica sa nižom konverzijom hrane češće bile zastupljene bakterije iz rodova Escherichia, Shigella i Salmonella.
Zanimljivi su rezultati Qi i sar. (Qi Z i sar., 2019), koji su analizirali strukturu i raznolikost cekalnog mikrobioma brojlera i nosilja i to sekvenciranjem segmenta bakterijskog dela ribozoma (16S rRNK). Rezultati ukazuju da su bakterije iz grupe Firmicutes, Bacteroidetes i Proteobacteria bile najčešće detektovane i kod brojlera i kod nosilja. Međutim, kada analiziramo bakterije na nivou roda, onda su nosilje imale češće prisustvo bakterija iz sledećih rodova: Bacteroides, Rikenellaceae, Phascolarctobacterium, Desulfovibrio, Prevotellaceae, kao i neklasifikovanih predstavnika bakterija Bacteroidales, dok je kod brojlerskih pilića bilo zabeleženo veće prisustvo bakterija iz sledećih rodova: Alistipes, Rikenella, Ruminococcaceae i Lachnoclostridium. Primetno je da su bakterije iz roda Desulfovibrio bile mnogo češće prisutne u cekumima nosilja nego u cekumima brojlera.
Ukupno 59 bakterijskih rodova je bilo identifikovano u cekalnom mikrobiomu živine, uključujući i 39 rodova koji pripadaju kolu (phylum) Firmicutes, dok su 3 najdominantnije prisutna roda (Ruminococcus, Clostridium i Eubacterium) činila 5 procenata, a ostatak rodova je predstavljao do 1 procenat kola (phylum) Firmicutes (We i S i sar., 2013). Bacterioidetes je bio drugi najprisutniji phylum (kolo) bakterija i bio je predstavljen sa 6 rodova, uključujući i rod Bacteroides koji je činio oko 40 procenata prisutne populacije.
Korisna uloga intestinalnog mikrobioma živine na organizam domaćina (živine)
Kao što se vidi, svi delovi intestinalnog trakta živine su gusto naseljeni brojnim i složenim zajednicama mikroorganizama (bakterije, gljivice, arheje, protozoa i virusi) među kojima dominiraju bakterije. Mikrobiom creva živine formira jednu vrstu zaštitne barijere koja se sastoji od mikroorganizama "zalepljenih" za zid ćelija epitela (enterocite) koji tako onemogućavaju kolonizaciju creva sa drugim, a posebno patogenim bakterijama. (Yegani M i Korver DR, 2008).
Te iste bakterije mikrobioma proizvode vitamine (na primer vitamin K i vitamine B kompleksa), kratkolančane masne kiseline (sirćetna, buterna i propionska kiselina), organske kiseline (mlečna kiselina), antimikrobne komponente (bakteriocin), niže trigliceride i ujedno obezbeđuju indukciju imunskog odgovora (nespecifičnog) koji doprinosi odgovarajućoj ishrani i zaštiti domaćina (Jeurissen SH i sar., 2002, Yegani M i Korver DR, 2008, Dibner JJ i Richards JD, 2005, Apajalahti J, 2005).
Sa druge strane, taj isti crevni mikrobiom može predstavljati i izvor bakterijskih patogena kao što su bakterije vrsta Salmonella i Campylobacter, koje se mogu preneti na ljude ili učestvovati u daljem prenošenju rezistencije na antibiotike i time predstavljati ozbiljnu pretnju po javno zdravlje (Kumar S i sar., 2018, Mancabelli L i sar., 2016, Zhou W i sar., 2012).
Uobičajeni crevni mikrobiom živine može biti koristan, ali i štetan po domaćina. Neka istraživanja su dokazala da je ekstenzivno odgajana živina, manje osetljiva na patogene bakterije ako se uporedi sa istom vrstom životinja koje su odgajane u sterilnim uslovima (engl. germfree animals) (Koopman JP i sar., 1984).
Takođe je već dobro poznato da mikrobiom živine ima značajnu ulogu u stimulisanju razvoja imunskog sistema koji podrazumeva formiranje sloja mukoze, epitela, imunskih ćelija crevnog trakta (citotoksične ćelije, pomoćne T ćelije, ćelije koje sintetišu imunoglobuline i fagocitne ćelije), kao i ćelije same laminae propriae (Dibner JJ i Richards JD, 2005, Shakouri MD i sar., 2009, Oakley BB i sar., 2014). Sva nabrojana tkiva grade barijeru između domaćina i mikroorganizama omogućavajući borbu protiv neželjenih mikroorganizama crevnog trakta. U donjim partijama creva (cekumi i kolon) mikrobiom proizvodi energiju i hranljive sastojke kao što su vitamini, aminokiseline i kratkolančane masne kiseline (engl. short chain fatty acids – SCFA) i to od nesvarenih ostataka hrane, koji na taj način postaju ponovo iskoristivi za domaćina. Upravo kratkolančane masne kiseline dokazano imaju bakteriostatske osobine i sposobnost eliminacije nekih patogenih mikroorganizama kao što su bakterije Salmonella spp. (Ricke SC, 2003). Pored toga, kratkolančane masne kiseline predstavljaju izvor energije za životinju i stimulišu proliferaciju epitelnih ćelija creva, što uvećava resorptivnu površinu crevnog epitela i time olakšava iskorišćavanje hrane. Uz to, kratkolančane masne kiseline snižavaju pH vrednost u kolonu, što inhibira rast mnogih patogenih bakterija (Christl SU i sar., 1997).
Mikrobiom creva živine ima značajnu ulogu i u metabolizmu azotnih jedinjenja. Na primer, cekalne bakterije konvertuju mokraćnu kiselinu u amonijak koji ptice onda mogu da resorbuju i dalje koriste u sintezi aminokiselina kao što je glutamin (Vispo C i Karasov WH, 1997). I na kraju, deo azota iz hrane, bakterije koriste za izgradnju sopstvenih proteina koje nakon toga domaćin koristi kao izvor proteina ili aminokiselina (Metges CC, 2000).
Procenjuje se da oko 20 procenata gena u metagenomu crevnog mikrobioma živine kontroliše metabolizam ugljenih hidrata, oko 10 procenata kontroliše metabolizam proteina i aminokiselina, a 1–2 procenta su odgovorni za metabolizam masti (Qu A i sar., 2008). Proces fermentacije polisaharida igra važnu ulogu u cekumima živine i kolonu ljudi, kao i kod svih monogastričnih životinja, a taj proces ne utiče samo na zdravstveni status creva, već i na zdravstveno stanje celog tela (Stanley D i sar., 2014). Metaboliti proizvedeni od mikrobioma creva se svrstavaju u 3 glavne grupe produkata: nutriente (hranljivi sastojci) koji se koriste od strane organizma domaćina, uključujući tu i crevni epitel. Oni promovišu rast i poboljšavaju funkciju kratkolančanih masnih kiselina (SCFA), uključujući sirćetnu kiselinu, propionsku i buternu kiselinu, zatim biološki aktivne komponente (uglavnom vitamini B kompleksa) i štetne i toksične supstance koje predstavljaju rizik po zdravlje domaćina. Ovi proizvodi ispoljavaju lokalne efekte na konkurentske grupe bakterija u crevnom epitelu i crevnom imunom sistemu (engl. GALT – gutassociated lymphoid tissue, u prevodu limfoidno tkivo povezano sa crevima), isto kao i sa sistemskim efektima na metabolizam domaćina. Lokalni efekti uključuju inhibiciju konkurentske patogene mikroflore, zaštitu crevne mukoze i stimulaciju odgovora GALTa. Sirćetna kiselina (acetat) ima ključnu ulogu kao jedan od važnih supstrata glavnine metaboličkih reakcija, dok buterna kiselina predstavlja svojevrstan biomarker zdravlja creva.
Štetna uloga intestinalnog mikrobioma živine na organizam domaćina
U gornjim partijama creva (želudac i tanka creva) mikroorganizmi mikrobioma se takmiče sa organizmom domaćina za izvore proteina i energije koji dolaze iz konzumirane hrane i time konkurišu domaćinu. Međutim, u gornjim i u donjim partijama creva živine, mikroorganizmi stvaraju toksične metabolite (produkte katabolizma aminokiselina) i katabolizuju žučne kiseline, što sve izaziva depresiju rasta i smanjuje svarljivost masti.
Prisustvo mikrobioma creva živine dovodi do uvećane sekrecije mucina od strane mukoznog sloja creva, zatim do bržeg obnavljanja površinskog sloja epitelnih ćelija creva, sa ciljem kontinuiranog očuvanja vlažnosti gastrointestinalnog trakta kako bi se preventiralo vezivanje mikroorganizama za epitelne ćelije i njihov prodor u organizam domaćina. Usled toga je imunski sistem razvijeniji i luči više imunoglobulina klase IgA, koji imaju sposobnost specifičnog vezivanja za površinske epitope bakterija, regulišući na taj način optimalni bakterijski sastav u crevima (samo komensali) (Metges CC, 2000, Mitchell MA i Moretó M, 2006). Iako su ovi procesi u svojoj suštini korisni za organizam domaćina, oni ipak povećavaju potrebe domaćina za energijom i proteinima i samim tim utiču negativno na njegove proizvodne performanse.
Poremećaj ravnoteže mikrobioma creva se često označava i kao disbioza. Disbioza može biti definisana kao kvalitativni ili kvantitativni disbalans normalnog mikrobioma tankih creva koji može dovesti do posledičnih promena u digestivnom traktu kao što su: poremećaj funkcije crevne barijere (istanjen zid creva) i smanjena svarljivost hrane. Samim tim nastaje i uvećan rizik od prodora bakterija u organizam domaćina i upalnih reakcija (Teirlynck E i sar., 2011).
Krajnji produkti bakterijske fermentacije proteina su amonijak, fenol, indol i amini, koji ugrožavaju zdravlje creva (Millet S i sar., 2010). Crevne bakterije kao što su Clostridium, Enterococci i Bacteroides oslanjaju se na proteine kao izvor energije, naročito kada ugljeni hidrati nisu dostupni u dovoljnoj količini u sadržaju creva. Zbog toga hrana koja sadrži visok procenat proteina može stimulisati potencijalne patogene i protivupalni razvoj mikrobioma (Yao CK i sar., 2016).
Do pomenutog stanja disbakterioze mogu dovesti stresogeni faktori koji mogu biti neinfektivne i infektivne prirode. Neinfektivni stresogeni faktori uključuju uticaj životne sredine, neizbalansiranu ishranu, promene u ishrani, prisustvo mikotoksina, loše i neadekvatno upravljanje odgojem živine, kao i poremećaje u genetici domaćina (Teirlynck E i sar., 2011).
Infektivni faktori uključuju razne virusne, bakterijske i parazitske infekcije ili prisustvo toksičnih metabolita koji su proizvedeni od strane štetnih mikroorganizama kao što je Clostridium perfringens.
Mikrobiom živine i imunski sistem
Kao jedan od esencijalnih mukoznih organa imuniteta domaćina, creva obavljaju dva važna zadatka: resorpciju hranljivih sastojaka i odbranu od patogenih mikroorganizama. Intersticijalni imuni sistem uključuje u svoj sastav: robustan mukozni sloj, očuvan integritet epitelnih ćelija (IEC), izlučeni rastvorljivi imunoglobulin klase A (IgA) i antimikrobne peptide (AMPs).
Već je dokazano da mikrobiom creva živine ima važnu ulogu u očuvanju normalne fiziološke homeostaze, moduliranju imunskog sistema domaćina i uticaj na razvoj i metabolizam organa. Ta interakcija se najbolje može razumeti ako uporedimo razlike koje nastaju u izgledu i funkciji creva kod normalno odgajanih brojlerskih pilića na farmi (kvalitetniji profil mucina i veći broj "goblet" ili peharastih ćelija koje luče mucin) u odnosu na piliće odgajane u izolatoru (bez ikakvog prisustva mikroorganizama), pri čemu je jedina razlika među njima postojanje i nepostojanje mikrobioma. Kod istih pilića je ustanovljena i razlika u broju i vrstama receptora na T limfocitima. Crevni mikrobiom modulira i odgovor B ćelija, kao i produkciju IgA. Stoga se smatra da uticaj mikrobioma creva preko pravilnog razvoja imunskog sistema domaćina ima potencijalne implikacije i na optimalno zdravlje i produktivnost životinja tako što dovodi do uštede energije, smanjenja "naprezanja imuniteta" i pojave inflamacije. Mora se naglasiti da svaka modifikacija mikrobioma utiče na zdravlje i metabolički status domaćina, što zauzvrat utiče na mikrobiom. Na taj način su domaćin i mikrobiom ujedinjeni i "osuđeni" na večnu simbiozu.
Da ponovimo, opisana pionirska istraživanja "normalnih" brojlerskih pilića u komparaciji sa "bezmikrobnim" (engl. germfree) pilićima su nedvosmisleno dokazala da je izloženost mikroorganizmima neophodna i preko potrebna kako bi se odigrao odgovarajući razvoj i sazrevanje imunološkog sistema creva živine (Dibner JJ i sar., 1998). Bezmikrobne životinje su osetljivije na crevne infekcije, pokazuju smanjenu prokrvljenost (vaskularizaciju), smanjenu aktivnost enzima varenja, istanjenost mišićnog zida creva, smanjenu produkciju citokina i manji nivo imunoglobulina IgA u serumu, smanjenje veličine Pajerovih ploča i manji broj intraepitelijalnih limfocita (Shanahan F, 2002).
Crevni mikrobiom reguliše i produkciju antimikrobnih peptida (AMP) koje luče intestinalne epitelne ćelije u koje spadaju defenzini kao što su: lektin C tipa, ribonukleaze, angiopoetin 4 i S100 proteini koji brzo ubijaju ili inaktivišu sve mikroorganizme (Gallo RL i Hooper LV, 2012).
Još pre oko 40 godina, Nurmi i Rantala (Nurmi E i Rantala M, 1973) su uveli termin konkurentsko isključivanje (engl. competitive exclusion) opisujući smanjenje kolonizacije Salmonella spp. kod ptica kod kojih je oralnim putem unet crevni sadržaj (feces) od ptica slobodnih od Salmonella vrsta i to odmah po izleganju (prvog dana života). Konkurentsko isključivanje (ili suzbijanje) podrazumeva bitno smanjenu mogućnost kolonizacije patogenih mikroorganizama i to razmatramo kao posledicu nekoliko mogućih mehanizama: fizičko zauzimanje mesta, kompeticije oko resursa u "fizičkoj ili hemijskoj niši" ili direktnim fizičkim ili hemijskim dejstvom na potencijalnog kolonizatora (patogenu bakteriju). Zapaženo je da veoma nizak nivo Salmonella vrsta može kolonizovati crevni trakt mladih brojlerskih pilića, dok su starije ptice skoro rezistentne na tu infekciju. To je dovelo do pokušaja transfera zajednice mikroorganizama od odraslih zdravih jedinki na jednodnevne piliće, što je rezultiralo očekivanim povećanjem rezistencije na kolonizaciju vrstama Salmonella.
Slični eksperimenti (ali i tretmani) se izvode i kod ljudi kada je u pitanju uporno prisustvo infekcija sa patogenom C. difficile. Uprkos još uvek nedovoljnom poznavanju mehanizama koji dovode do ovakvih rezultata, najefikasniji pristup za prevenciju intestinalne kolonizacije živine sa Salmonella vrstama ostaje i dalje upravo navedeni tretman sadržajem creva poreklom od zdravih ptica. Svi pokušaji da se definiše tačno određen sadržaj creva ili da se proizvede mešavina određenih mikroorganizama nisu imali ni približno istu efikasnost kao sadržaj creva zdrave živine. Eksperimenti ovog tipa se u živinarstvu izvode još od 1965. godine (Schefferle HE, 1965). Kada bi mehanizmi dejstva bili otkriveni verovatno bi novi, potpuno "prirodan i zdrav" pristup profilaksi mnogih bolesti bio na dohvat ruke.
Gastrointestinalna zajednica mikroba se ponaša kao antiinfektivna barijera vršeći inhibiciju adhezije patogena i sprečavanjem njegove kolonizacije. Ovo se realizuje proizvodnjom bakteriocina i drugih toksičnih metabolita za patogene, dok se fermentiranjem kompleksnih polisaharida iz hrane obezbeđuje iskoristiva energija za domaćina. Takođe, početne interakcije između crevne zajednice mikroorganizama i urođenog imunološkog sistema domaćina mogu dovesti do adaptivnog (stečenog) imunskog odgovora koji može biti zavisan od B ili T ćelija (Pan D i Yu Z, 2014). Mnogi faktori mogu uticati na uspostavljanje i evoluciju mikrobioma gastrointestinalnog trakta, ali interakcije između mikrobioma creva i imunskog sistema su očigledno dominantne u nekim stadijumima njegovog razvoja. Infekcije gastrointestinalnog trakta koje su detektovane od strane imunskog sistema domaćina (živine) izazivaju neki od odgovara preko složenog sistema povezivanja puteva koji uključuje i urođene i stečene imunološke odgovore i reakcije.
Betadefenzini su mali antimikrobni peptidi koji se nalaze na površini epitela creva i čine važan deo urođenog imuniteta (Shimizu M i sar., 2008) Ovi peptidi su proizvedeni kao odgovor na prisustvo lipopolisaharida koji uključuju i TLR4 (engl. Tolllike receptors) i faktore transkripcije NFκB (Akbari MR i sar., 2008). Na ekspresiju defenzina u crevima utiče izloženost creva mikroorganizmima i zato redukcija u izloženosti mikrobiomu rezultira mnogo slabijom ekspresijom duodenalnog i cekalnog betadefenzina u ranom periodu rasta ptice (Butler VL i sar., 2016). Treba dodati da zajednica mikroba pomaže u isključivanju patogenih mikroba, fermentiranju nekoliko supstrata, kao i pružanju energije domaćinu (Pan D i Yu Z, 2014).
Nastaviće se u sledećem broju
Literatura je dostupna kod autora milankosekler@yahoo.com